 Gatherer,
D. G.
(2004). The Inherent Instability of Memetic Systems:Use of a Genetic
Algorithm to Solve a Parameter Optimisation Problem in a Memetic
Simulation .
Gatherer,
D. G.
(2004). The Inherent Instability of Memetic Systems:Use of a Genetic
Algorithm to Solve a Parameter Optimisation Problem in a Memetic
Simulation .
Journal of Memetics - Evolutionary Models of Information Transmission,
8.
http://cfpm.org/jom-emit/2004/vol8/gatherer_d.html
The Inherent Instability of Memetic Systems:
Use of a Genetic Algorithm to Solve
a Parameter Optimisation Problem in a Memetic Simulation
Derek Gatherer
dgatherer@talk21.com
Abstract:
'Memetic isolation' is a situation in
which a society or culture exhibits a cultural trait not found in any
neighbouring cultures (
Gatherer 2002b).
A previously developed simulation, consisting of a grid of connected
societies of cultural agents, is further analysed to define the
circumstances under which memetic isolation is maximised and minimized.
Parameters varied include migration, and cultural interaction within
and
between societies. Some stereotypical societies are simulated, and the
differences in outcomes are analysed statistically. A genetic algorithm
is then used to discover the combinations of parameters that produce
extreme results. Minimization of memetic isolation appears to be simply
a matter of cultural or natural selection on the meme concerned. By
contrast, maximization of memetic isolation requires an intuitively
unlikely combination of low cultural interaction, high migration and no
selection. The classic memetic theoretical result, that horizontally
transmitted memes tend to be more spatially clustered than vertically
transmitted memes or genes, is shown to depend on the existence of
cultural bridges, or 'weak ties', between societies and also to be at
the mercy of migration rates and selective forces.
Key words:
Contagion, evolutionary epidemiology of culture, cultural evolution,
cultural selection, meme, allomeme, cultural trait, genetic algorithm,
globalisation.
1. Introduction
A large literature exists on the relationship
between genetic isolation and cultural isolation (e.g. Sokal et al. 1989;Barbujani & Sokal 1990; Vona et al. 1996;
De Silvestri &
Guglielmino 2000; Gonzalez
Jose et al. 2002). Genetic isolation may be the result of a
'founder effect', where a population expands rapidly from a small set
of
original progenitors. This often happens in colonial situations, e.g.
in South Africa (Torrington
& Viljoen 1991), Venezuela (Ramirez-Duque et al.
1982; Vivenes De
Lugo et al. 2003), Canada (Scriver
2001) or Finland (Kittles
et
al. 1999). It may also follow a sudden 'population crash'
caused by a natural disaster or epidemic disease, followed by
regeneration of the population from the survivors (Ambrose 1998; Harpending et al. 1998).
All of these factors may also cause cultural isolation, which can then
contribute to further genetic isolation if interbreeding with
geographically neighbouring populations is prevented or reduced by
cultural factors. However, the two phenomena are not necessarily
related, and one may exist without the other.
Cultural isolation may be seen as an extreme case of
'memetic isolation' (Gatherer 2002b),
defined here as the situation in which any cultural trait of a society
is different to the corresponding mutually exclusive cultural trait,
the
'allomeme' (Durham 1991), of any of its
surrounding cultures. A true cultural isolate will be memetically
isolated for an extensive selection of cultural traits.
In classic mathematical memetic theory, 'vertical
transmission' refers to
traits
that are passed on from one generation to the next, specifically from
parents to progeny (a variant is 'oblique transmission' in which traits
are passed within families, but not necessarily from parent to child).
By contrast 'horizontal transmission' refers to transmission within a single
generation, either between siblings or other genetic relatives, or
within a non-related peer group (Cavalli-Sforza &
Feldman 1981). One of the results of this body of theory is that
traits with more than one means of transmission have a greater tendency
to homogeneity within populations, and also that horizontally
transmitted
traits are more likely to be spatially clustered (Cavalli-Sforza &
Feldman 1973, Uyenoyamaet
al.. 1979, reviewed by Cavalli-Sforza
1979). 'Spatial clustering' in this context means that individuals
exhibiting the trait tend to be found in closer geographical proximity
than those who do not exhibit the trait.
The present paper further investigates this area of
theory using agent-based simulation. The simulation was previously
described in detail in Gatherer (2002b),
and may be considered as a variant of the Social
Interaction Model (SIM) simulation of Axelrod (1997).
Two questions are asked:
1)
Under what combination of parameters is memetic isolation maximised?
2)
Under what combination of parameters is memetic isolation minimised?
The answers provided enable us to refine the
predictions of the original theoretical treatments of transmission and
its relation to spatial clustering of traits (reviewed by Cavalli-Sforza 1979).
2. The Model
Since frequent comparison is made to the Social
Interaction Model simulation of Axelrod (1997),
this is described first. Imagine a set of distinct societies or
cultures arrayed in geographical space. Let each culture consist of a
small number of cultural traits, in the order of 10 or so. Adjacent
cultures may interact if they have a sufficient number of cultural
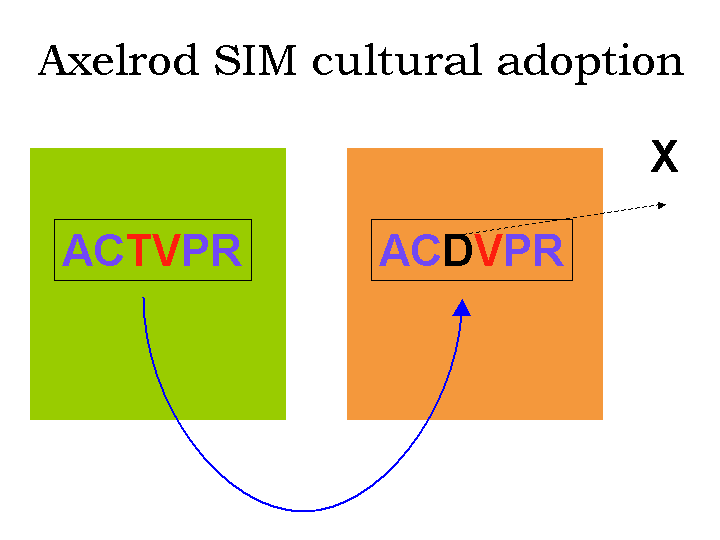
traits in common. This situation is illustrated in Figure
1, where a minimum of four out of six cultural traits in common are
required in order for the societies to interact with each other.
Figure 1:
Rules for possibility of interaction in Axelrod's SIM.
Cultural traits are represented by letters, and coloured to show which
are found in common between pairs. Since the upper pair has 4 out of 6
cultural traits in common, these two societies can interact. However,
the lower pair only has 2 traits in common and therefore cannot
interact.
Once the two societies have established an interaction, one society
may randomly adopt a cultural trait from the other. This is illustrated
in Figure 2. At the end of the interaction,
one of the societies has become slightly more similar to its neighbour.
It is easy to see how this simple situation mirrors the process of
cultural transfer in real societies. Cultures with very differing
languages, religions or habits may fail to make the necessary cultural
contact. However, once a certain threshold has been crossed and
communication is established, cultural interaction can commence. This
model makes the assumption that there is no selection for or against
any
cultural trait, or that traits are imposed wholesale, e.g. by conquest.
Figure 2:
Adoption of one cultural trait from adjacent society in interacting SIM pair. Trait X in the right society is replaced by
trait V from the left society. Having had 4 traits in common, the pair
of societies now has 5 in common.
This process is repeated over several cultural generations in a grid
of several cultures, typically 10 by 10, starting from a random
distribution of traits. One of the principal results of SIM
is that cultural 'blocs' are formed, i.e. areas on the cultural
landscape with several near-identical cultures. This is especially the
case if the probability of interaction is varied according to the
relatedness of a pair of societies, i.e. if a pair of societies with 5
traits in common has more chance of interaction that a pair with only 3
in common, and so on. Intuitively, it can be seen that his tends to
mean
that societies that have previously interacted will be increasingly
likely to interact further in future generations, thus producing a
positive feedback.
The variation on SIM
presented previously by Gatherer (2002b)
uses an identical concept of a grid of societies in geographical space.
The size of grid presented in the scenarios below is 10 x 10, the same
order of magnitude used by Axelrod (1997).
However, the focus is switched to the individual agent with the society
rather than the society as a whole. SIM
assumes that each society is culturally homogeneous, insofar as each
society has a single set of cultural traits. Figure 3 shows how this
may
be adapted to allow individuals within societies to differ in their
repertoire of memes and genes. The word 'genes' in this case is used as
a shorthand for 'strictly vertically transmitted memes', since in
haploid organisms (i.e. those with a single copy of each gene in their
genomes) these categories are formally equivalent (Cavalli-Sforza &
Feldman 1981). In panel A
of Figure 3, a grid of nine societies may be
seen.
Individuals are indicated as circles with their vertical memetic
component as the letter A or B, and their horizontal memetic
component as the colour red or blue. These individuals may perform
three kinds of lifecycle activity: reproduction, migration and cultural
interaction. In Figure 3A and 3C, cultural interaction is
indicated with a black arrow if it occurs within societies (e.g. in the
bottom right society of panel A,
an individual with blue meme teaches it to an individual with red meme,
thus resulting in that individual having blue meme in panel B), and a green arrow if it occurs
across inter-societal boundaries. Reproduction is indicated with a
white
arrow and the newly produced individuals are surrounded by a dotted
line
in 3A and 3C. Migration is indicated by a blue
arrow, and can only occur to neighbouring societies (in other words
this
is a 'two-dimensional stepping stone' model of migration - Rychkov & Sheremetyeva
1977). Panels 3B and 3D illustrate the end result after
the
processes displayed in 3A and 3C. The societies in panels A and B are coloured blue or orange
according to the majority of blue or red memes respectively. In panels C and D the societies are coloured yellow
or grey according to the majority of genes (or strictly vertical memes)
A and B, respectively.
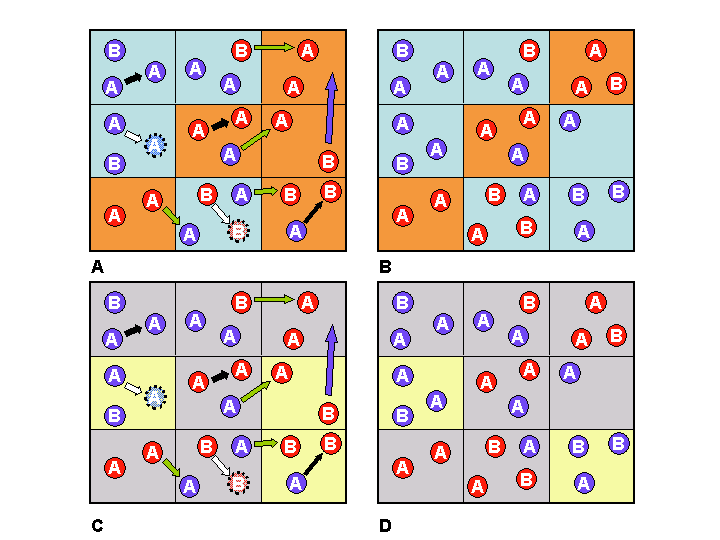
Figure 3:
Reproduced from Gatherer (2002b). A and B labels are strictly vertical
memes,
red and blue colours are horizontal memes. Thus a blue individual
labelled B has the B vertical meme and the blue
horizontal meme. Black arrows: horizontal cultural interaction within
societies. Green arrows: horizontal cultural interaction between
societies. Blue arrows: migration of individuals. White arrows:
reproduction (new progeny are identical to their parents and are shown
with dotted outlines). Orange squares: societies with majority red
memes. Light blue squares: societies with majority blue memes. Grey
squares: societies with majority of A
vertical memes/genes. Yellow squares: societies with majority of B vertical memes/genes. Fuller
explanation given in text.
In summary, the simulation consists of a grid of connected societies
populated by cultural agents, its dynamics being defined with a set of
four simple parameters:
- the rate of population growth, r
- the rate at which teaching/learning
occurs, o
- the rate at which teaching/learning
occurs between societies, n
- the rate at which individuals migrate
to neighbouring societies, m
These parameters are designated mnemonically as r, o, n and m, respectively
(i.e. reproduce, own-teach,neighbour-teach and migrate). All take
values
between zero and one, and represent probabilities that are either
user-defined or created using a random number generator. The parameters
n and o are
interdependent, with o always
evaluated first, and then n evaluated
depending on the result of the evaluation of o. For example
when o is
0.5, and n
is 0.6, an agent will exhibit successful cultural transmission during
its lifetime at probability 0.5. If that agent does exhibit cultural
transmission, there is then a probability of 0.6 that that cultural
transmission will be to a member of a neighbouring society, and 0.4
that
it will be to a member of the same society. This could be re-expressed
in terms of the notion of 'strong' and 'weak' ties, where o is the
probability of existence of a tie, and n the
probability that such a
tie
is 'weak' (sensu Granovetter 1973). A pseudocode representation of the
process is as follows:
INITIALIZATION:
For each of the 100 cells in the 10-by-10 array:
{
place two Agents in each cell
randomly assign one of four Gene attributes to each individual
randomly assign one of four Meme attributes to each individual
}
ITERATION:
For 100 generations do the following:
{
For each Agent:
{
If Agent Meme attribute = "A" and CultSel = "Y"
{
Double parameters n and o for that agent
}
If Agent Meme attribute = "A" and NatSel = "Y"
{
Double parameter r for that agent
}
If a random number x < r
{
Reproduce Agent
}
If a different random number y < o
{
If same random number y is also < n
{
Transmit Meme to any Agent in any adjacent cell
}
else
{
Transmit Meme to any Agent in same cell
}
}
If a different random number z < m
{
Migrate Agent to adjacent cell
}
}
}
END.
Additionally, 'cultural selection' or 'natural
selection' may be applied to one of the memes in the population. Here,
these terms are used in the sense of Cavalli-Sforza & Feldman (1981), as follows: a
culturally selected meme is one that has an increased probability of
transmission relative to its allomemes; a naturally selected meme is
one
that provides a survival or reproductive advantage to those individuals
that exhibit that behaviour or cultural trait.
Figure 4 provides a
diagrammatic representation of the agent in UML (Universal
Modelling Language). The simplicity of this agent (all attributes and
methods public, no constructors etc) is due to its implementation as a
Perl record rather than a true object in Java or C++. Perl records are
fairly analogous to struct in C. Agents have one of four Meme
attributes
("A", "B", "C",
"D") and one of four Gene
attributes ("1", "2", "3",
"4").
|
Agent
|
|
+Gene: String
+Meme: String
+Location: Vector
|
|
+Reproduce(): Agent
+Teach_Own(): Agent
+Teach_Neighbour(): Agent
+Migrate(): Agent
+Die()
|
Figure 4: UML
diagram for the Agent class. Attributes are above the line and Methods
below. Public scope is represented by a "+" (all are public)
Perl scripts are provided in the Appendix that:
- run the simulation with graphical output
- implement a genetic algorithm (GA) to optimise the parameters.
Both scripts require Perl, and the first also
requires the Tk graphics library. A recent version of ActivePerl (Build
6 series or more recent, freely available from http://www.activestate.com)
provides all the necessary library components. All scripts provided
have been tested on RedHat Linux 6 running Gnome, and on Microsoft
Windows 98.
3. The Effects of Varying
Parameters on Memetic Isolation
The 'memetic state' of a society for a certain
cultural trait, is here defined by the 'first-past-the-post' voting
method. The Meme attributes of each agent in the society are scored,
and
if allomeme "A" scores more than any of the other three Meme
attributes,
the memetic state of that society is "A".
If allomeme "A" is not the
highest scoring attribute, the memetic state of that society is "not-A". 'Memetic isolation' occurs when
that society's 'memetic state' is "A"
and that of all its immediate neighbours is "not-A". The overall memetic isolation
is the percentage of societies that are in this state. It is admitted
that 'first-past-the-post' is potentially a flawed means of scoring a
society, but it is often used in sociological contexts. For instance,
statements such as the "Belgium is a Catholic country" are simply
expressions of such head counts. Qualifying those situations where the
'first-past-the-post' voting produced a narrow majority, perhaps by the
use of a 'marginal' label, could further refine the model. However,
this would be complex enough to merit another paper, and so is not
dealt
with here.
'Genetic isolation' can similarly be calculated
using the Gene attribute "1".
Since the agents are haploid (i.e. they have only a single Gene
attribute), the Gene is equivalent in terms of its transmission to a
strictly vertically transmitted meme, as demonstrated by Cavalli-Sforza
& Feldman (1973).
It is thus possible to contrast 'memetic isolation' and 'genetic
isolation' in terms of the likelihood of isolation for horizontal and
vertical memes. Therefore, 'genetic isolation' is used hereafter simply
as a shorthand for 'memetic isolation for strictly vertical traits'.
3.1 Scenario 1: The Random Background -
the "strictly vertical model"
The scenario is initialised with two individuals per
cell, and allowed to run for 100 generations with a rate of population
growth (r)
of 0.01 per generation. Migration (m), and the two
social interaction variables (o and n) are set at
zero. This simulates a situation where societies grow slowly in size,
but there is no 'horizontal
transmission' (Cavalli-Sforza
& Feldman 1981), or movement of individuals across society
boundaries. It is essentially a non-cultural population, or perhaps
more
exactly a population where culture is contained strictly within
vertical
lineages. This is given merely to demonstrate the properties of the
system in a background state, where as little as possible is happening
except the effects of randomness over a large number of cycles. It is
difficult to think of a real-world example of this kind of cultural
situation; even non-human species with aspects of culture, such as song
birds and great apes, do not limit cultural transmission within genetic
lineages (e.g. Burnell 1998;Lynch et al. 1989; Reader & Laland 2000; van Schaik et al. 2003).
The scenario was repeated 100 times and the average memetic and genetic
isolation measured. These were found to be 3.23% and 3.00%
respectively.
Statistical significance was assessed using a paired t-test. In this
and
all subsequent scenarios, t-tests are two-tailed, as there is no a
priori expectation that parameters will increase or decrease levels of
isolation. A paired t-test between the genetic and memetic isolation
values for each of the 100 runs, was not significant, thus indicating
that both horizontal and vertical memes behave identically in strictly
vertical populations. In effect, the 'horizontal memes' are forced to
become vertical memes by the absence of cultural interaction in the
population, and thus this scenario constitutes a sort of reductio ad
absurdum. However, it is necessary to demonstrate the basic background
behaviour against which other scenarios will be measured.
3.2 Scenario 2: Introducing Cultural
Exchange Within Societies - the "desert
island model".
Social interaction within societies is now
permitted, by raising parameter o to 0.5. This
means that each agent will transmit a horizontal meme with probability
0.5 within its lifetime, in other words half of all agents transmit
horizontally per generation, and half do not. All other parameters are
the same. This recreates the situation of societies between which there
is no contact, but which have a flourishing social life of their own,
such as might occur on extremely isolated oceanic islands. It should be
noted at this point, and for all subsequent scenarios, that it is
recognised that real societies are infinitely more complex than any
simulation can reproduce. The purpose of adding labels such as 'desert
island' to the simulations, is really to serve as a mnemonic shorthand,
and also to perhaps act as an 'intuition pump' for practical
applications.
After 100 generations, over 100 runs of the
scenario, the average genetic and memetic isolation values are 3.16%
and
4.08% respectively. A paired t-test shows these to be statistically
significantly different at p<0.0004. Thus, against a background of
no
migration and no cultural exchange between societies, social
interaction
within our artificial societies increases the likelihood that those
societies will be memetic isolates. This is because the rate of the
random walk amongst competing allomemes in those societies is speeded
(see Gatherer 2002a, Fig. 2). If o is raised to
1,
allowing each agent a successful cultural transmission event per
lifetime, the figures are essentially unchanged. This is called a
'desert island' model, since it would apply only to societies that were
completely cut off from the rest of the outside world. Some examples
are known, of which the best studied is perhaps that of the Bounty
mutineers on Pitcairn Island, who remained completely undiscovered for
decades after settling, and in effective cultural isolation for some
years after. In this period they were able to evolve a unique culture
incorporating both European and Polynesian elements, including a new
creole English language (Refshauge
& Walsh 1981). If one imagines a thought experiment in which
the Bounty mutineers colonised two separate islands, mutually
unconnected, the model predicts that they would become memetic isolates
for many traits. This rarely applies to many island archipelagos, as
the
inhabitants are often in contact, albeit infrequently. This leads to
the
next scenario where such an arrangement is permitted.
3.3 Scenario 3: Introducing Limited
Cultural Exchange Between Societies - the "Palaeolithic
model".
Parameter n is now raised
from zero to 0.01. This permits cultural exchange with a neighbouring
society at a probability of 0.01 x o per individual
per generation. Therefore at o of 0.5, the
rate of inter-cultural exchange is 0.005. The societies are now no
longer totally isolated "desert islands", but have the kind of casual
contact perhaps normal for sparsely distributed and occasionally
trading
nomadic bands. Over 100 runs of the scenario, again for the standard
100 generations, average genetic isolation remains around the
background
level at 3.12%, but memetic isolation drops to 1.78%. A paired t-test
shows this to be a significantly different value for memetic isolation
at levels of p tending to zero. Thus even a small amount of cultural
exchange between our artificial societies may, ceteris paribus, create
tendencies to cultural homogenisation on a local geographical scale.
Cultural blocs (Axelrod 1997) thus begin
to emerge out of a patchwork
background. Where parameter n is increased,
memetic isolation falls even further: at n = 0.1, memetic
isolation falls to 0.71%. Thus in a scenario of 100 societies, most
runs terminate without a single memetic isolate at the end of 100
generations. This is called the 'Palaeolithic model' as it very roughly
recreates a situation hypothesised to have existed in Late Ice Age
Europe, where nomadic bands of hunters of large herds of bison,
mammoths
etc, would have existed in isolation punctuated by brief seasonal
periods of contact at assembly points, perhaps associated with exchange
of gifts, arrangement of marriages and ritual behaviour (e.g. Rudgley
2000). A more modern example may be provided by the Andamanese, who
remained in relative isolation within their archipelago for many
thousands of years (e.g. Thangaraj
et al. 2003). Again it bears
reemphasis that the correspondence between the artificial societies and
the anthropological examples given is quite naïve, and really just
raises the issue of how better models of such societies could be
created, rather than claiming to have created one here.
3.4 Scenario 4: Introducing Natural
Selection - the "Neolithic model".
Meme "A" is
now submitted to natural selection. This means that those agents with
Meme attribute "A" have twice
the normal chance of reproduction of those with the other three
allomemes. Meme "A" agents thus
reproduce above replacement at a probability of 0.02 per generation,
instead of the 0.01 for the other three allomemes. This is termed here
the "Neolithic Model" as it is reminiscent of one of the main theories
for the Neolithic transition in Europe, namely that the cultural
activity of farming provided a reproductive and survival advantage, in
that early farmers were able to feed more children and live longer to
reproduce more, compared to their Mesolithic hunter-gatherer neighbours
(Renfrew 1987). If m, n and o are all set to
zero, so that there is no migration or cultural exchange within the
population, but natural selection is permitted to increase r from 0.01 to
0.02 for Meme "A" individuals,
genetic isolation after 100 generations is 3.68% and 2.34%. This
memetic
isolation is significantly different to the background "pre-cultural
model" (See 3.1), although both models have no cultural exchange. The
reason for this is that the evolutionary advantage of Meme "A" in terms of natural selection
means that this meme will tend to take over the populations in which it
is found. If it happens to occur in two adjacent societies, even at
initial low frequencies, it will eventually run to fixation in those
societies. Of course, this is not really a good "Neolithic model", as
Neolithic societies also had cultural exchange as well as natural
selection for a meme (assuming for the present that this particular
theory of Neolithic transition by demic expansion is true). A second
"Neolithic model", allowing levels of cultural exchange equivalent to
the "Palaeolithic model", i.e. o = 0.5 and n = 0.01, has
levels of genetic and memetic isolation of 3.37% and 1.11%,
respectively. A t-test shows that this is a significantly different
level to that when there is no natural selection. Natural selection of
a
meme thus reduces its likelihood of memetic isolation.
3.5 Scenario 5: Introducing Cultural
Selection - the "Contagion model".
The second "Neolithic model" above, now has natural
selection of Meme "A" replaced
by cultural selection. This means that Meme "A" will be transmitted at double the
normal frequency of cultural exchange. Thus with o = 0.5 and n = 0.01, as
above, Meme "A" has
probabilities of transmission of oA = 1 and nA = 0.02 . Genetic and memetic
isolation after 100 generations average to 3.24% and 0.21%,
respectively. Therefore, cultural selection almost removes memetic
isolation entirely.
3.6 Summary of scenarios
Table 1 show the summary of
the
scenarios
|
Scenario number, number of section where
described and name of model
|
m
|
n
|
o
|
select.
|
Genetic isolation (%)
|
Memetic isolation (%)
|
|
1: 3.1 Strictly vertical
|
0
|
0
|
0
|
None
|
3.23
|
3.00
|
|
2: 3.2 Desert island
|
0
|
0
|
0.5
|
None
|
3.16
|
4.08
|
|
2: 3.2 Desert island - 2
|
0
|
0
|
1
|
None
|
3.26
|
4.04
|
|
3: 3.3 "Palaeolithic"
|
0
|
0.01
|
1
|
None
|
3.12
|
1.78
|
|
3: 3.3 "Palaeolithic" - 2
|
0
|
0.1
|
1
|
None
|
3.33
|
0.71
|
|
4: 3.4 "Neolithic"
|
0
|
0
|
0
|
Natural
|
3.68
|
2.34
|
|
4: 3.4 "Neolithic" - 2
|
0
|
0.01
|
0.5
|
Natural
|
3.37
|
1.11
|
|
5: 3.5 Contagion
|
0
|
0.01
|
0.5
|
Cultural
|
3.24
|
0.21
|
Table 1:
Summary of the average genetic and memetic isolations resulting from
the various scenarios. The 'strictly vertical' model provides the
random background level of isolation. The 'desert island model' shows
the highest level of memetic isolation. m: migration, n: cultural
interaction within societies, o: cultural
interaction between societies
Several questions may now be answered by using
t-tests to compare the statistical significance of differences between
rows in Table 1. In this section, t-tests are
two-tailed and non-paired, since comparison is between different
scenarios. An F-test is first carried out to establish if the t-test
should be heteroscedastic or homoscedastic.
What factors affect genetic isolation?: Method:
t-tests between genetic isolation data for all rows against row 3.1
(where there is the random background level of genetic isolation).
Answer: no model significantly changes the likely degree of genetic
isolation of each society. The differences within the range of 3.12% to
3.68% are all explicable as statistical random variation. This applies
equally to cultural traits that are strictly vertically transmitted.
What factors affect memetic isolation?: Method:
t-tests between memetic isolation data for all rows against row 3.1
(where there is the random background level of memetic isolation).
Answer: by contrast with the above, all models have a level of memetic
isolation significantly different to the random background at
p<0.01.
The answer is therefore completely different to the above, in that one
might say that several factors may influence memetic isolation,
including cultural and natural selection, and the rates of cultural
transmission.
Does the "Neolithic - 2" have less memetic isolation
than the basic "Neolithic" model? Method: t-tests between memetic
isolation data for rows "3.4 Neolithic" and "3.4 Neolithic - 2".
Answer:
yes, at p tending to zero. Therefore, it is possible to conclude that
an increase in social interaction within a society decreases memetic
isolation when the meme is under natural selection, whereas it
increases
it when natural selection is not operating (compare "Strictly vertical"
and "Desert Island").
Does "Contagion" have less memetic isolation than
any of the other models? Method: t-tests between memetic isolation data
for all rows against row 3.5. Answer: yes, "Contagion" has a
significantly lower level of memetic isolation than all other models at
p tending to zero. It is thus possible to say that culturally selected
traits are unlikely to exist in isolation.
4. The Genetic Algorithm
The above set of simulations only dealt with seven
scenarios as compared to a random background. However, since there is a
combinatorial explosion of possible scenarios, a heuristic method must
be used to select the scenario most likely to give rise to extreme
conditions.
A standard method for use in such situations is the Genetic Algorithm (GA - reviewed by Bentley 1999). The general form of the GA used here is as follows:
START: Randomise representations of the parameters in a linear string.
FOR n CYCLES
{
Assess the result of those parameters by 100-generation simulation
Rank strings by the result under investigation (e.g. maximal memetic isolation)
Discard very worst performers
Recombine and/or mutate remainder of unacceptable perfumers
Retain and reproduce acceptable performers to maintain constant numbers of strings
}
END.
In the GA
presented here, the threshold below which GA
strings are discarded is 0.775 standard deviations below or above the
average memetic isolation, depending on whether the GA
is attempting to maximise or minimise memetic isolation. The threshold
for mutation and/or recombination is 0.5 standard deviations
above/below
the average. Mutation and recombination occur with probabilities of
0.05
and 0.1 respectively, but only within the sub-standard group. The
better performing strings are thus protected from mutation and
recombination (this is quite a Lamarckian GA!).
These properties were derived by a process of empirical trial and
error, and there are many other forms of GA
that could be used. The balance between homogenisation, due to
selection, and diversity, due to mutation and recombination, is a
delicate one, and has to be solved afresh for every situation in which
a GA
is used. It should be noted that the fitness landscape is extremely
noisy. In all the scenarios above in section 3,
the standard deviation of the memetic isolation is high. For this
reason, it appears to be more profitable to protect the better
performing strings. The GA
process is illustrated in diagrammatic form in Figure
5, and the continuing high standard deviation may be seen in Figure 6.
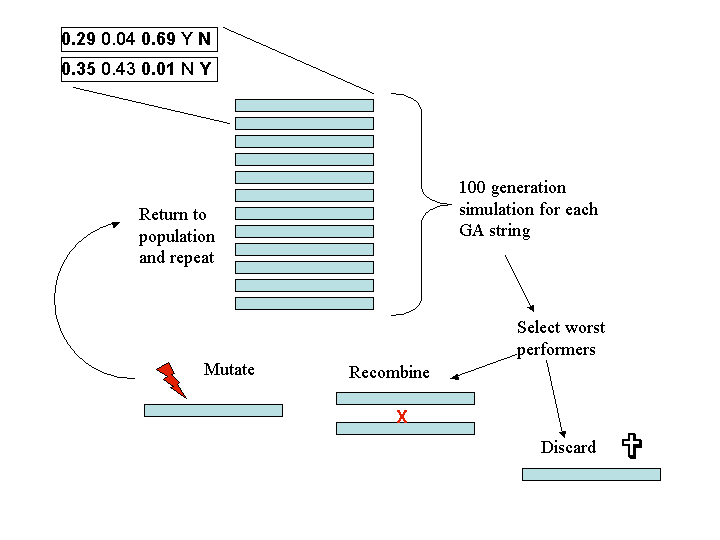
Figure 5:
Diagrammatic representation of the GA process. The
starting sets of parameters for the model are represented as a string
of
length 5. Each string is run as a separate simulation, and the results
are assessed according to the desired outcome (e.g. high memetic
isolation or low memetic isolation). The 'worst' performers according
to
the desired criteria are either discarded, mutated or recombined with a
randomly chosen other member of the population. The discarded strings
are replaced by randomly chosen strings from the better performing part
of the population. Selection of GA strings thus tends to homogenise the
population while mutation and recombination maintain some variability.
The whole process was repeated over 100 cycles.
A set of GA
strings was prepared by semi-randomly initialising the variables, m, n and o, and the
selective conditions for cultural and natural selection, 50 times. The
initialisation is semi-random as the starting set was biased to give
fairly low levels of migration. The initial set produced is shown in
abbreviated form in Table 2. After 100
iterations of a GA
process, selecting for maximum memetic isolation at each stage, the 50
populations corresponding to the 50 GA
strings have average memetic isolation of 4.60% and average genetic
isolation of 4.72%. The set after 100 GA
generations is shown in abbreviated form in Table 3.
The most obvious feature of the action of the GA
over 100 generations of searching for best parameters for maximum
memetic isolation, is to eliminate cultural and natural selection. The GA
also optimises memetic isolation by reducing the rate of cultural
exchange within the culture, variable o, from the
initial average of 0.462 to less than 0.005. Migration is increased
from
0.048 at initialisation to 0.559. The optimal value for teaching to
other societies is also maintained fairly high, at 0.432 from 0.501 at
initialisation, but this will still represent a small level of cultural
exchange since the background probability of cultural exchange is less
than 0.005.
|
Growth rate
|
Teach own society
|
Teach other society
|
Migration
|
Cultural Selection
|
Natural Selection
|
|
1 0.01
|
0.24
|
0.89
|
0.04
|
Y
|
N
|
|
2 0.01
|
0.41
|
0.46
|
0.07
|
Y
|
Y
|
|
3 0.01
|
0.44
|
0.58
|
0.05
|
N
|
Y
|
|
Several rows........
|
|
49 0.01
|
0.12
|
0.06
|
0.01
|
N
|
N
|
|
50 0.01
|
0.14
|
0.90
|
0.07
|
Y
|
N
|
|
Average
|
0.462
|
0.501
|
0.048
|
21 Y
|
18 Y
|
Table 2: Truncated listing of 50 GA strings semi-randomly initialised at 5 parameters.
|
Growth rate
|
Teach own society
|
Teach other society
|
Migration
|
Cultural Selection
|
Natural Selection
|
|
1 0.01
|
0
|
0.81
|
0.5
|
N
|
N
|
|
2 0.01
|
0.01
|
0.81
|
0.5
|
N
|
N
|
|
3 0.01
|
0
|
0.81
|
0.5
|
N
|
N
|
|
Several rows........
|
|
49 0.01
|
0
|
0.81
|
0.5
|
N
|
N
|
|
50 0.01
|
0
|
0.81
|
0.5
|
N
|
N
|
|
Average
|
0.0048
|
0.432
|
0.5592
|
0 Y
|
0 Y
|
Table 3:
Truncated listing of 50 GA
strings after 100 generations of the GA
process, selecting for maximum memetic isolation.
When the same process is performed, but the
selective conditions are to minimise memetic isolation, the resulting
average parameters are compared in Figure 4.
The minimisation GA
was only performed over 7 rounds of selection, as compared to 100
rounds
for maximization, as it rapidly achieves a near-zero rate of memetic
isolation.
|
GA seln. (rounds)
|
Teach own soc.
|
Teach others
|
Mig.
|
Cult. Seln.
|
Nat. Seln.
|
Mem. Isol.
|
Gen. Isol.
|
|
Minimise (7)
|
0.49
|
0.60
|
0.49
|
23/25 Y
|
23/25 Y
|
0.02%
|
3.43%
|
|
Maximise (100)
|
0.0048
|
0.432
|
0.5592
|
0/25 Y
|
0/25 Y
|
4.60%
|
4.72%
|
Table
4: Comparison of 25 rounds of
GA
selection for the optimal parameters to maximise memetic isolation,
with
7 rounds of selection for the optimal parameters to minimise memetic
isolation.
The progress of the memetic isolation-maximising GA
is shown in Figure 6.
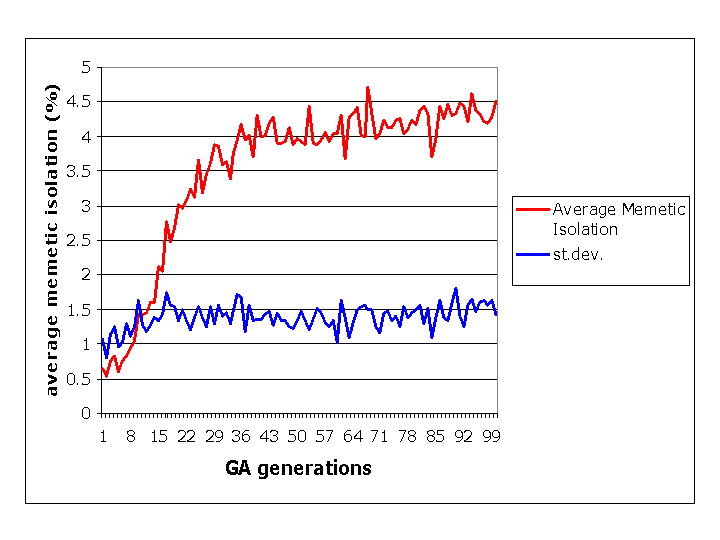
Figure 6: Progress of the genetic
algorithm in selecting parameters to maximise memetic isolation. Red:
the average memetic isolation in the 50 populations corresponding to
the
50 GA
strings. Blue: the standard deviation of that average throughout the 50
populations. The persistence of a relatively high standard deviation
illustrates the 'noisiness' of the system.
5. Discussion
The contemporary
phenomenon of globalisation involves cultural as well as economic
aspects. It has been estimated that 5.6% of the world's languages are
in
danger of extinction, and a further 4.4% have recently become extinct (Sutherland 2003). In the Social
Interaction Model (Axelrod 1997), this
phenomenon was seen as an emergent property. SIM is
another grid-based agent model in which neighbouring agents are
permitted to exchange information with other societies providing they
shared at least one cultural trait. SIM
has a single agent per grid square, which can either be taken as an
individual or as a whole, and necessarily homogeneous, society (Axelrod
suggests 'homogeneous villages'). There is no migration. SIM
leads to the increasing homogenisation of agents in close proximity,
while at the same time agents that had by chance no common traits with
their neighbours, remain completely differentiated. If the SIM
agents are treated as individuals, SIM
demonstrates how neighbouring individuals can form homogeneous cultural
areas. Similarly, if the SIM
agents are regarded as homogeneous societies, SIM
demonstrates the formation of cultural 'blocs' on a local geographical
basis. The model presented here differs from SIM,
in that it is necessarily based on individuals. Societies are simply
geographical areas, whose cultural profiles depend on those of the
agents that inhabit them. Another important difference is that in the
present model, the level of social interaction between societies is
constant for each run of the scenario, and is defined by parameter n. However, in SIM,
the analogous parameter is proportional to the number of cultural
traits that neighbouring agents have in common. Thus, if there are no
traits in common, neighbouring agents will not interact, thus creating
a
local 'desert island' model. If the SIM
agents are seen as homogeneous societies, SIM
thus simultaneously operates a 'desert island' model and a
'Palaeolithic
model' within the same scenario, with the two models operating in
different parts of the grid, with shifting boundaries as the scenario
progresses. A model more similar to the present one in its structure,
is
given by Ray et al. (2003),
but those authors focus exclusively on the genetic phenomena and do not
deal with horizontally transmitted cultural entities.
The genetic algorithm
process presented here seeks to derive parameters for the prevention of
cultural blocs. It delivers maximal memetic isolation (an average of
4.6% of societies are memetic isolates after 100 generations of the
scenario) at levels of social interaction tending to zero (less than
0.5% per person per lifetime), and with high migration (over 55% per
person per lifetime), where no selective conditions operate. This
describes a rather nightmarish scenario where there is little or no
social communication between continually wandering individuals. There
is some superficial similarity here to theories of 'alienation' (Geyer 1994). Although technically, this might
be the optimal way of maintaining memetic diversity in a grid of
artificial societies, it is clearly not a solution that any would
prescribe in the real world. It is also completely vulnerable to
selection pressures where they arise. Even in situations of very low
social interaction, a culturally selected meme will tend to spread
through a population (see Gatherer 2002a,
Fig. 4). When there is no social interaction at all, and cultural
selection ceases to operate, natural selection can still push memes to
fixation through growth of the lineages that exhibit them. In effect,
extreme migration rates cause the cellular nature of the scenario to
break down, and effectively become a single society. Parameters o and n then cease to
have any significance, as cultural transfer occurs principally by
transfer of people.
How would one make the
model more comparable to the real world? Empirical studies of
Australian Aborigines suggest a realistic figure of 0.12 to 0.13 for m, where the
pattern of migration is mostly to neighbouring tribal groups and rarely
to more distant tribes, reflected in the present model (Lasker &
Crews 1996). Migration rates of m ~ 0.55 are
rare
in the real world. Where they do occur they are often in specific
directions, e.g. invasions, flight of refugees, economic migration, and
are not reproducible in the present model. Even when they do, they are
often smaller than m
~ 0.55. For instance even at the height of the potato famine in
Ireland, the total decline in population over the decade 1841-1851 was
about 20%, including migration and death (although of course a decade
does not constitute a lifetime, so the figures are not strictly
comparable). The largest migration occurring in a single year, the 17
million people that migrated from one part of the Indian subcontinent
to
another after partition, still only accounted for some 7% of the Indian
population at the time. Modern radiochemistry is beginning to show that
Neolithic populations may have been more mobile than previously thought
(e.g. Montgomery et al.
2000). There are some interesting questions that could be
addressed in this area using scenarios of the sort presented here, once
accurate parameters can be estimated from the empirical data, such as
why late Neolithic Malta became a cultural isolate after two millennia
in contact with the rest of the Mediterranean (e.g. Robb 2001; Schulting & Richards 2002).
Empirical studies to determine realistic values of parameter o may be
more problematic, but there may be some clues from anthropological
field
studies of cultural transmission (e.g. Hewlett &
Cavalli-Sforza 1986).
When set to minimise
memetic isolation, the GA simply identifies cultural
selection as the over-riding factor in this. As modelled here,
contagious memes run to fixation within societies, and when transferred
to neighbouring societies, do the same there. However, it is also clear
that this is a naïve representation of the likely course of
contagion within a society. Gatherer (2002a)
shows that there are several factors that can derail 'the contagionist
paradigm', and that random, or pseudo-random, walks are more likely
than
sigmoid curves when considering meme frequencies within any society.
The effect of modern
mass communication media is to amplify the horizontal mode of
transmission so that one individual can communicate with many more
individuals simultaneously than would be possible in a lifetime of
individual contacts (Cavalli-Sforza
& Feldman 1981). This situation is not explicit modelled
either here, or in SIM, but could easily be
incorporated. Axelrod (1997) suggests a
model where long-range interactions are permitted and probabilities of
interaction are based on such long-range communication networks, rather
than geography. Gould & Tornqvist (1971) describe a transition from
a spatially contagious pattern of cultural spread to a hierarchical one
dominated by rapid transfers between urban centres. These authors cite
Pyle's (1969) work on cholera by way of
example: the 1832 epidemic in Montreal and New York spread slowly down
the Ohio/Great Lakes river system. In 1865 by contrast, several major
cities, by that time connected by rail, were struck simultaneously, and
the second wave of the epidemic struck towns connected to those major
cities. Thus, for societies in the industrial phase of development, it
is clear that such a spatial grid model (whether using SIM
or the model presented here) is inadequate.
However, although the
initial model was conceived as a representation of a geographical grid
of societies, in order to compare the results of simulations with a
geographically ordered ethnographic data set (Gatherer
2002b), it is also possible to consider it more abstractly, as a
single large society partitioned into smaller social networks. This
effect is in any case forced on the scenario at high values of m, such as that
produced by the GA.
The arrangement of the cells on the grid does not then refer to any
geographical separation of sub-societies, but to their separation in
terms of the likelihood of weak ties. Sub-societies that are not
adjacent on the grid will have no weak ties at all between them, and
those that are adjacent will have weak ties with probability n per individual
per generation.
In summary, it has been
known for over a quarter of a century that memes have a greater
tendency
to homogeneity within populations than genes, and also that memes are
more likely to be spatially clustered (reviewed by Cavalli-Sforza 1979). In the
context of the present scenario, 'spatial clustering' can be taken to
mean an absence of memetic isolation, or in Axelrod's terms the
formation of 'cultural blocs'. The scenarios presented here demonstrate
that, where societies are totally isolated (the "desert island" model, section 3.2) the tendency to memetic isolation
is actually increased. However, where cultural exchange, even at a tiny
level, is permitted between neighbouring societies, memetic isolation
declines precipitately (the "Palaeolithic" model, section
3.3). Where the meme is also under selection, especially cultural
selection, memetic isolation falls towards zero (the "Neolithic" and
"Contagion" models, sections 3.4 and 3.5). Therefore the inherent tendency of memes
to be more spatially clustered than genes depends on the existence of
cultural bridges, or 'weak ties' (Granovetter
1973), between societies. The GA results
confirms
the primacy of selective pressures in creating spatial homogeneity, but
also suggests that such homogeneity can be rapidly fractured by high
migration rates. Memetic systems can be seen to be highly unstable,
especially when the selective forces at work within them are also
cultural. Culture is always close to chaos.
Appendix: Code Listings
All listings require Perl to be installed, and should run under both
windows and Unix. In the event of any problems, please contact
the
author.
The code for the
graphical simulator:
The code for the GA:
References
Ambrose SH (1998) Late
Pleistocene human population bottlenecks, volcanic winter, and
differentiation of modern humans. Journal
of Human Evolution 34,
623-651.
Axelrod R (1997) The
dissemination of culture: a model with local convergence and global
polarization. Journal of Conflict
Resolution 41, 203-226.
Barbujani G
& Sokal RR (1990) Zones of sharp genetic change in Europe are also
linguistic boundaries. Proceedings
of
the National Academy of Sciences, USA 87,1816-1819.
Bentley P (1999) (ed.).
Evolutionary Design by Computers.
Morgan Kaufmann, San Francisco.
Burnell K (1998)
Cultural variation in savannah sparrow, Passerculus sandwichensis,
songs: an analysis using the meme concept. Animal Behaviour 56, 995-1003.
Cavalli-Sforza
LL & Feldman MW (1973) Models for cultural inheritance. I. Group
mean and within group variation.. Theoretical
Population Biology 4,
42-55.
Cavalli-Sforza
LL & Feldman MW (1981) Cultural
Transmission and Evolution. Princeton University Press,
Princeton.
Cavalli-Sforza
LL (1979) Cultural change and its relevance for human genetics. Ciba Foundation Symposium 66, 5-23.
De
Silvestri A & Guglielmino CR (2000) Ethnicity and malaria affect
surname distribution in Consenza Province (Italy). Human Biology 72, 573-583.
Durham WH (1991) Coevolution: Genes, Culture and Human
Diversity. Stanford University Press, Stanford.
Gatherer D (2002a)
The Spread of Irrational Behaviours by Contagion: An Agent
Micro-Simulation. Journal of
Memetics
- Evolutionary Models of Information Transmission 6. http://cfpm.org/jom-emit/2002/vol6/gatherer_d.html
Gatherer D (2002b)
Identifying Cases of Social Contagion Using Memetic Isolation:
Comparison of the Dynamics of a Multisociety Simulation with an
Ethnographic Data Set. Journal of
Artificial Societies and Social Simulation 5(4). http://jasss.soc.surrey.ac.uk/5/4/5.html
Geyer F (1994)
Alienation, Participation and Increasing Societal Complexity. Kybernetes 23, 1-3
Gonzalez
Jose R, Garcia-Moro C, Dahinten S & Hernandez M (2002) Origin of
Fueguian-Patagonians: an approach to population history and structure
using R matrix and matrix permutation methods. American Journal of Human Biology 14, 308-320.
Gould P
& Tornqvist G (1971) Information, Innovation and acceptance. In:
Hagerstrand T & Kuklinski A (eds.) Information
Systems for Regional Development, Lund Series in Geography, Ser.
B, Human Geography 37, 149-168.
Granovetter M
(1973) The strength of weak ties. American
Journal of Sociology 78,
1360-1380.
Harpending
HC, & al. (1998) Genetic traces of ancient demography. Proceedings of the National Academy of
Sciences, USA, 95,
1961-1967.
Hewlett
BS & Cavalli-Sforza LL (1986) Cultural transmission among Aka
pygmies. American Anthropologist 88, 922-934.
Kittles
RA, & al. (1999) Autosomal, mitochondrial, and Y chromosome DNA
variation in Finland: evidence for a male-specific bottleneck. American Journal of Physical Anthropology
108, 381-399.
Lasker GW
& Crews DE. (1996) Behavioral influences on the evolution of human
genetic diversity. Molecular
Phylogenetics and Evolution 5,
232-240.
Lynch
A, Plunkett GM, Raker A & Jenkins P (1989) A model of cultural
evolution of chaffinch song derived with the meme concept. The American Naturalist 133, 634-653.
Montgomery
J, Budd P & Evans J (2000) Reconstructing the lifetime movements of
ancient people: a Neolithic case study from southern England. European Journal of Archaeology 3, 370-386.
Pyle G (1969) Diffusion of
cholera in the United States. Geographical
Analysis 1, 59-74.
Ramirez-Duque
P, Arends T & Merino F (1982) Chediak-Higashi syndrome: description
of a cluster in a Venezuelan-Andean isolated region. Journal of Medicine 13, 431-451.
Ray
N, Currat M & Excoffier L (2003) Intra-Deme Molecular Diversity in
Spatially Expanding Populations. Molecular
Biology and Evolution 20,
76-86.
Reader SM
& Laland KN (2000) Diffusion of foraging innovations in the guppy. Animal Behaviour 60, 175-180.
Refshauge
WF & Walsh RJ (1981) Pitcairn Island: fertility and population
growth, 1790-1856. Annals of Human
Biology 8, 303-312.
Renfrew C (1987) Archaeology & Language: The Puzzle of
the Indo-European Origins. Jonathan Cape, London.
Robb J (2001) Island
identities: ritual, travel and the creation of difference in Neolithic
Malta. European Journal of
Archaeology 4, 175-202.
Rudgley R (2000) Secrets of the Stone Age: A Prehistoric
Journey. Century, London.
Rychkov
YG & Sheremetyeva VA (1977) The genetic process in the system of
ancient human isolates in North Asia. In: Harrison GA (ed.), Population Structure and Human Variation,
Cambridge University Press, Cambridge, 47-108.
Schulting
RJ & Richards MP (2002) The wet, the wild and the domesticated: the
Mesolithic-Neolithic transition on the west coast of Scotland. European Journal of Archaeology 5, 147-189.
Scriver CR (2001)
Human genetics: lessons from Quebec populations. Annual Review of Genomics and Human
Genetics 2, 69-101.
Sokal
RR, Oden NL, Legendre P, Fortin MJ, Kim JY & Vaudor A (1989)
Genetic differences among language families in Europe. American Journal of Physical Anthropology 79, 489-502.
Sutherland WJ
(2003) Parallel extinction risk and global distribution of languages
and
species. Nature 423, 276-279.
Thangaraj
K, & al. (2003) Genetic Affinities of the Andaman Islanders, a
Vanishing Human Population. Current
Biology 13, 86-93.
Torrington
M & Viljoen DL (1991) Founder effect in 20 Afrikaner kindreds with
pseudoxanthoma elasticum. South
African Medical Journal 79,
7-11.
Uyenoyama
M, Feldman MW & Cavalli-Sforza LL (1979) Evolutionary effects of
contagious and familial transmission. Proceedings
of the National Academy of Sciences, USA 76, 420-424.
Van
Schaik CP, & al. (2003) Orangutan cultures and the evolution of
material culture. Science 299, 102-105.
Vivenes
De Lugo M, Rodriguez-Larralde A & Castro De Guerra D. (2003)
Beta-globin gene cluster haplotypes as evidence of African gene flow to
the northeastern coast of Venezuela. American
Journal of Human Biology 15,
29-37.
Vona
G, & al. (1996) Genetics, geography, and culture: the population of
S. Pietro Island (Sardinia, Italy). American
Journal of Physical Anthropology 100, 461-471.
(c)
JoM-EMIT 2004