Journal of Memetics - Evolutionary Models of Information Transmission, 6.
http://cfpm.org/jom-emit/2002/vol6/castro_l&toro_ma.html
We suggest that human cultural learning was made possible by the simultaneous appearance, in one of our hominid ancestors, of two capacities: the capacity to imitate others behaviour and the capacity to approve or disapprove of others behaviour. With the help of a mathematical model, we have studied the conditions that allow the evolution of both capacities. We consider four different genotypes: "the only-learner" that learns by trial and error, "the imitator" that learns by trial and error and imitation, "the only-assessor" that learns by trial and error but that can also approve or disapprove of offspring's behaviour and, finally, "the assessor", who behaves like the imitator but, he can approve or disapprove of offspring's behaviour. The assessor genotype is the best genotype and the only-learner genotype is the worst when the learned behaviour that would be culturally transmitted is adaptive. If this behaviour is maladaptive, the genotype only-assessor is the best genotype and the genotype assessor can be the worst genotype. Notwithstanding, in this situation, the assessor can also be better than the imitator and even better than the only-learner. The success of assessor is due to his capacity to increase the phenotypic correlation between parents and offspring, and thus speeding the rate at which natural selection, if present, will increase or decrease the frequency of learned behaviours.
Keywords: Conceptual categorisation; assessor; imitation; teaching; cultural evolution.
In species having a certain trial and error learning capacity the
development of a system of cultural transmission can turn out to be adaptive
because individuals do not need to reinvent behaviour. Boyd & Richerson
(1985) propose that cultural transmission provides a short-cut in time
and costs as compared to individual learning. However, culture has only
reached an important level of development in Homo sapiens. In non-human
animals forms of cultural transmission are absent or limited to a narrow
range of behaviours, as is the case of song learning in song birds, or
rudimentary, as seems to be the case in apes (Whiten & Ham, 1992; Heyes
& Galef, 1996). Besides, in recent years, monkeys imitative ability
has been questioned (Galef, 1988; Visalberghi & Fragaszy, 1990; Tomasello,
1996). At the heart of the controversy lies an interesting distinction
between "true imitation", in which one animal copies the behaviour pattern
of a performer it has observed, and "stimulus enhancement", in which a
performers action merely focuses an observers attention on critical environmental
features, increasing the speed with which the observer subsequently learns
a similar behaviour pattern through its own efforts. This can have evolutionary
importance, because cumulative culture change is possible when there is
imitation but not when there is stimulus enhancement (Boyd & Richerson,
1995). Several reviews suggest that all known cases of animal social traditions
can be explained as the result of stimulus enhancement (Tomasello, 1990;
Galef, 1992; Tomasello, 1996), although the question still remains open
(Whiten, 1998; Byrne & Russon, 1998; Custance et al., 1999; Whiten
et al., 1999).
In the literature on cultural transmission, trial and error learning and imitation are usually two alternative kinds of learning device (see Aoki, 2001, for a review). Cultural organisms can have in their brains a switching mechanism by which the organism can use either one or the other under different circumstances. In these models, each organism can learn individually or socially, but it cannot use both options at once. When individuals use trial and error learning, they learn the appropriate behaviour but they suffer learning costs. When they use imitation, individuals adopt permanently the behaviour of the individual that they use as a model and, therefore, avoid the costs of individual learning, but suffering a cost if they imitate an inappropriate model. Stable equilibrium might be achieved through a mixed strategy involving either polymorphism between imitative and creative individuals, or more realistically, individuals displaying both modes of behaviour (Boyd & Richerson, 1985; Feldman et. al., 1996; Henrich & Boyd, 1998).
In the model we propose here the relationship between trial and error learning and imitation is considered in a different manner. The adoption of a behaviour is defined as a process with three steps: the first, to discover and to learn the behaviour, the second, to evaluate behaviour as good or bad, and the third, to reject or to incorporate it into the behavioural repertory. Cultural organisms can to discover and learn a behaviour -the first step- both through trial and error learning or imitation. Thus, imitation increases the probability of discovering and learning a behaviour pattern from parents, but afterwards the individuals have to evaluate the learned behaviour through their evaluative brain structures and they can adopt or reject the behaviour that has been imitated. The same process occurs when cultural individuals discover by trial and error behaviours that they have not observed in other individuals. Therefore, imitation is an alternative process to trial and error learning with respect to discover a behaviour but not with respect to its final rejection or adoptation. We think that this model describes better the interaction between imitation and trial and error learning in the chimpanzées and it must be the starting point to study the evolution of cultural transmission in the hominids.
Moreover, we consider that our hominid ancestors have developed the conceptual capacity to categorise behaviour. This capacity is defined as the ability to categorise one's own and others' behaviour through a non verbal conceptual value code -positive/negative, good/bad- which works with the same evaluative brain structures that control trial and error learning (Castro, 1992; Castro & Toro, 1995; Castro & Toro, 1998). This represents the transformation of an automatic mechanism of categorizing behaviour in a mechanism of conceptual categorization. We have suggested that hominids with this ability to categorise -that we call hominid assessor- could approve or disapprove of other's behaviour. In this paper, we analyse how this ability to approve or disapprove of behaviour can be adaptive if it occurs between parents and offspring because parents can help the offspring to accept the good behaviour and to reject the bad one. Ethnographic records show teaching, defined as an instructed learning of behaviours, abilities or skills, are rare among human foragers and in most simple human societies (Rogoff, 1990; Tomasello et al., 1993; Tomasello, 1999). However, we think that the transmission from parents to offspring of information on the evaluation of the learned behaviour is a process of cultural transmission that must be denominated teaching and we suggest that teaching, defined in this way, is present in all human societies. That is, teaching is characteristic of our species.
Our model is a model of gene-culture evolution but we think like Kendal and Laland (2000) that these kind of models could be very useful in memetics. More specifically our aim is to emphasise the importance that the ability to approve or disapprove of others behaviour has for the meme evolution.
The adoption of a behaviour through trial and error learning can
be divided into three stages. The first is to discover and to experiment
with the behaviour. The second is to evaluate the behaviour, and the third,
to accept or to reject it. We will call "status" the set of behaviours
that an individual discovers and evaluates during his ontogeny.
The model assumes that behaviour can be described as a dichotomy: to do something (1) vs. to do nothing (0). If there is only one behaviour 1 for example, to eat a particular vegetable (1) vs. not to eat it (0)- the status of an individual can be status 1 or status 0 if he knows or does not know behaviour 1, respectively. The probability of an individual discovering and testing behaviour 1 is h -the probability of this behaviour being part of his status. In this model, behaviour 0 to do nothing- is always present in the status.
At the end of the ontogenic period, the individual has to decide the
behaviours that he finally will incorporate into his behavioural repertory.
If the status is 1, b is the probability that
he decides to incorporate behaviour 1 instead of rejecting it. The parameters
h
and b represent the probability of complex processes
that depend on many factors. We deliberately exclude the observation of
other individuals behaviour from those factors that affect parameters
h
and b .
All individuals can discover the behaviour through trial and error
learning, but cultural individuals can also discover it through imitation.
Imitation is introduced in the model by a parameter a
that represents the probability of learning a behaviour by observing it
in the parents -vertical cultural transmission. Imitation increases the
probability h that an individual learns and tests the active behaviour
that his parents exhibit but it does not affect the probability b
of accepting or rejecting it. For simplicity, we assume that cultural transmission
of behaviour is determined by choosing as a model only one of its parents,
the mother. The matrix of transmission between parents and children is
given in table 1.
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
where h* = (h + (1 - h) a ) is the probability of an individual learning the behaviour 1 when his mother is 1. In this expression, a represents the intensity of the process of cultural transmission and (1-h)a measures the net effect of this process. At the end of the period of ontogeny, the individual decides the behaviour that he finally will exhibit depending on the same parameter b as in the individual learning case.
The model assumes that only the active behaviour 1 can
be observed and, therefore, can be culturally transmitted, but not the
absence of a behaviour, 0. Therefore, 1 is a cultural trait but 0 is
not. In the discussion we consider the situation where both behaviours,
1 and 0, can be imitated. We also consider what happens when other individuals
different from parents are imitated -i.e., oblique and horizontal transmission..
The model assumes that some individuals that we call assessors have
the ability to approve or disapprove of learned behaviour of their offspring
whereas non-assessor individuals do not have this ability. The parameter
d
measures the intensity of approval or reproof made by the assessor parent
(0 £ d £ 1).
This parameter d increases or decreases the
probability b of offspring accepting a behaviour
depending on whether it has been approved or disapproved of by the parents.
The behaviour 1 will be approved if the parental model has adopted this
behaviour and it will be disapproved of only if the parental model knows
this behaviour -it is present in its status- but it has rejected it. For
simplicity, only one of the parents, the mother, approves or disapproves
of a child's behaviour. If the status is 1, b*
is the probability that assessor prefers to incorporate behaviour 1 instead
of rejecting it. b* depends on the evaluation
of the behaviour:
If the evaluation is positive b* will be
b* =b + (1 - b )d
If the evaluation is negative b* will be
b-* =
b-
bd
The process of transmission is assumed to be controlled by a sexual
haploid genetic system with two loci, one for imitation and the other for
categorisation, with two alleles per locus (I/i) and (A/a), respectively.
Therefore, four genotypes are distinguished:
- ia: "non-imitator and non-assessor" or "only-learner" (without ability either to imitate or to transmit evaluations):a = d = 0
- iA: "only-assessor" (with ability to transmit evaluations but without ability to imitate):
a = 0 and 0 < d < 1
- Ia: "imitator" (with ability to imitate but without ability to transmit evaluations):
0 < a < 1 and d = 0
- IA: "assessor": (with ability to both imitate and transmit evaluations):
0 < a < 1 and 0 < d < 1
Each one of these four genotypes can adopt three different phenotypes depending on the status and the behaviour exhibited and therefore there will be 12 distinct phenogenotypes. For example, for only-learner: ia-00 (they dont do 1 because they dont know 1), ia-01 (they dont do 1 because they dont want), and ia-11 (they do 1), where the superscript represents the status. It is also assumed that the only differences among genotypes are due to the values of a and d .
The use of a haploid genetic system is justified because it provides an approximation to the situation of intermediate dominance in diploid systems, especially near of the state of allelic fixation (Feldman & Cavalli-Sforza, 1984). Mating occurs randomly and from each mating the offspring genotypes are produced with the standard Mendelian probabilities, using the recombination fraction r. Mutation is assumed to be absent. The action of natural selection occurs when the individual has decided his behaviour after the ontogeny period and before mating. The fitness of phenotypes 0 and 1 is 1 and 1+s, respectively, s being the selective value. Each individual is given a certain probability of removal from the population based on s.
The possible costs of learning are not considered in the model, because we assume that these costs are the same across all the different genotypes. Obviously, this can be unrealistic. In fact, imitation allows the offspring to quickly get to know the behaviours that the parents exhibit and, if these behaviours are adaptive, they can profit from them earlier. However, this rapidity can be a disadvantage if the imitated behaviour is maladaptive for the offspring, either because it is harmful in itself or because the offspring is too young to practice the behaviour correctly. Furthermore, the capacity to imitate does not preclude the possibility that the offspring explores and evaluates not only the imitated behaviours but also other new behaviours that the parents do not exhibit and, therefore, the offspring also suffers learning costs. In the other hand, the capacity to approve or disapprove of offsprings behaviour can facilitate the evaluation of the behaviour, but it can also have a fitness cost c that represents a cost of parental care. We have preferred to ignore the possible differences in the learning costs and to analyse only the existence of fitness cost c for only-assessor and for assessor.
The evolution of phenotypic frequencies in the absence of selection,
together with the equilibrium values in genotypically monomorphic populations
and in the absence of selection is given in the Appendix. From there, it
can be seen that the equilibrium frequency of the phenotype 1 in these
populations follows the decreasing order
IA > Ia > iA = ia
The allele I allows imitation. The imitation decreases the frequency of status 0 and increases the frequency of status 1, and this results in a decrease in the equilibrium frequency of phenotype 0 and an increase in the equilibrium frequency of phenotype 1, the behaviour that can be imitated. The effect of imitation is stronger when the behaviour is more difficult to learn by individual learning -low values of h. In fact, if h = 1 there is no difference between imitator and only-learner. Obviously, the effect of imitation will increase as a , the ability to imitate maternal behaviour 1, increases. For a given value of h, the effect of a is stronger as bincreases.
The allele A allows their carriers, when they act as mothers, to approve
or disapprove of the behaviour that their offspring has learned if and
only if they know the behaviour, that is, if it is present in the mother
status. The A allele increases the phenotypic correlation between mothers
1 (11) and their offspring and between mothers 0 with status
1 (01) and their offspring. This phenotypic correlation will
increase as the parameter d -the ability to
transmit evaluations- increases. If d = 1, all
offspring of mothers 1 that acquire status 1 will be 1 and, on the contrary,
all the offspring of mothers 0 with status 1 (01) will be 0.
The effect of d is stronger as b
decreases.
With selection, the system is too complex to be solved analytically,
although it can be easily iterated in a computer (the software used for
simulation is available from the authors upon request). The frequency of
the different phenogenotypes when equilibrium is reached has been calculated
in different situations for populations composed of the four genotypes
with selection either in favour or against the phenotype 1 (figures 1-8).
The initial distribution of four genotypes was 25% for each one. The initial
distribution of status and phenotypes was 100% 00. The recombination
fraction r was 0.5, although using other values does not qualitatively
change the results.
With selection there is always a genotype that eliminates the others and goes to fixation. We can analyse the ranking among the four genotypes by looking at the order in which they are eliminated. If the behaviour that can be imitated 1 has positive selective value (figures 1-4), assessor is always the best genotype and only-learner is the worst. The other two genotypes, imitator and only-assessor have an intermediate position. From the frequency at equilibrium without selection of phenotype 1 in each one of the genotypically monomorphic populations we would expect that the relative fitness of genotypes would be Ia > iA, because the genotype imitator Ia produces a greater frequency of individuals 1 than genotype only-assessor iA (figures 1, 3, and 4). However, this is not always the case. The genotype only-assessor iA can be better than genotype imitator Ia if the increase in the frequency of phenotype 1 produced by imitation is small -h big-, the effect due to selection s is great enough, and the effect of the transmission of evaluations is great -b small (figure 2). Thus, only-assessor is advantageous in the probably uncommon situations where the individuals reject behaviours that are adaptive.
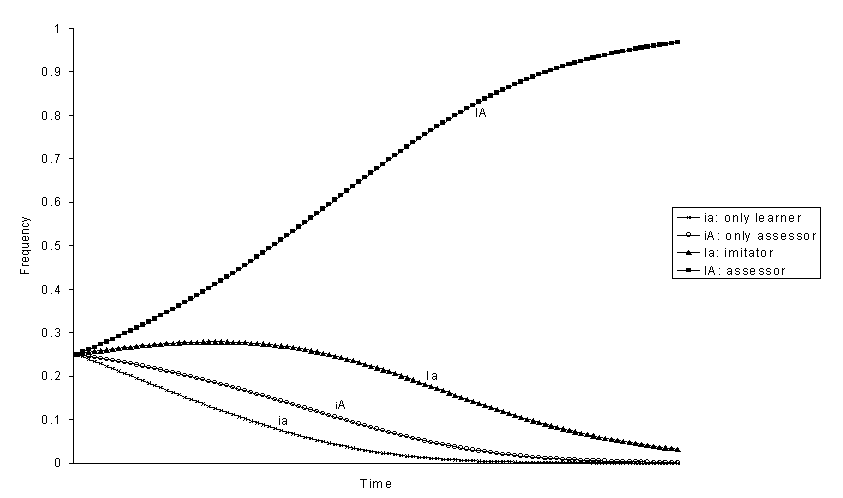
Figure 1. Evolution of genotypic frequencies with positive selection
for phenotype 1 ![]()

Figure 2. Evolution of genotypic frequencies with positive selection
for phenotype 1 ![]()
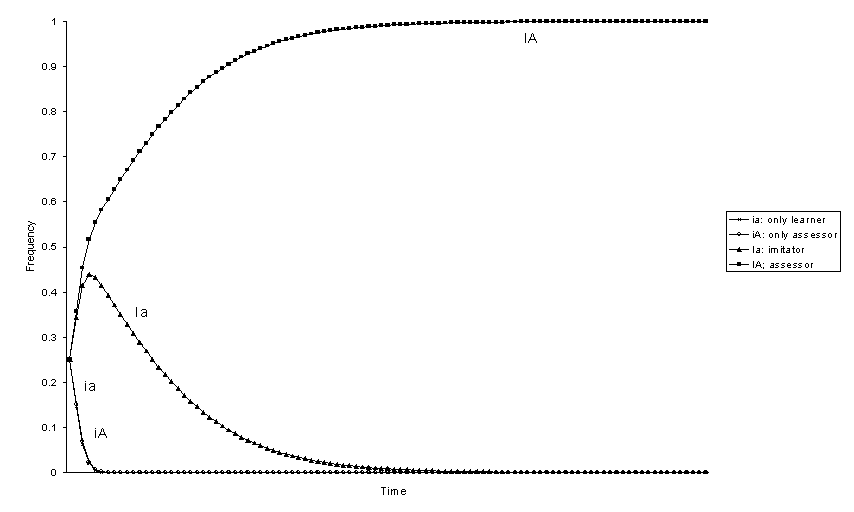
Figure 3. Evolution of genotypic frequencies with positive selection
for phenotype 1![]()
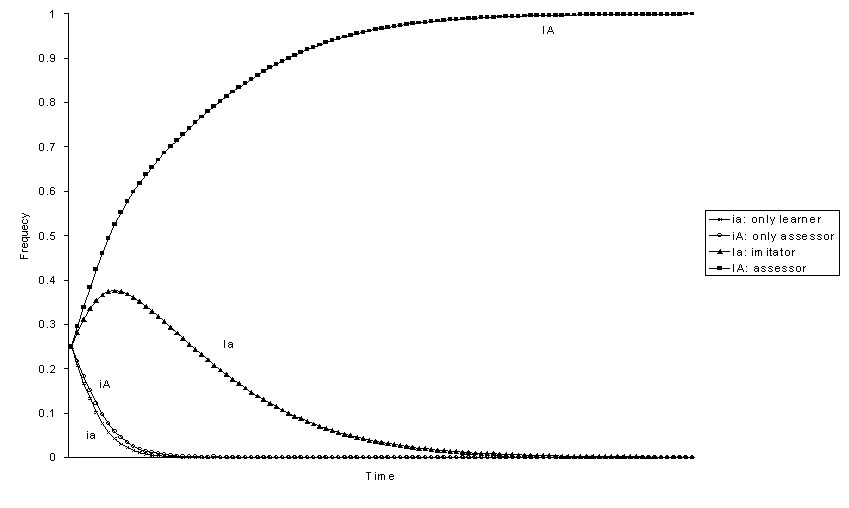
Figure 4. Evolution of genotypic frequencies with positive selection
for phenotype 1 ![]()
If behaviour that can be imitated 1 has negative selective value, only-assessor is always the best genotype and eliminates the others (figures 5-8). The reason why only-assessor is superior to only-learner under these circumstances is because only-assessors are able to increase the fitness of their offspring. From the frequencies at equilibrium without selection, the expected ranking of the genotypes should be: iA > ia> Ia > IA (figures 5 and 7). However, the genotype assessor can be better than imitator, iA > ia > IA > Ia (figure 8), if the selection coefficient s against phenotype 1 is great and the effect of imitation increasing the frequency of behaviour 1 is small h big. And it can even be better than only-learner iA > IA > ia > Ia (figure 6), if the probability b of accepting the maladaptive behaviour 1 is sufficiently small. Thus, assessor is advantageous if the maladaptive behaviour is easy to learn and to reject.
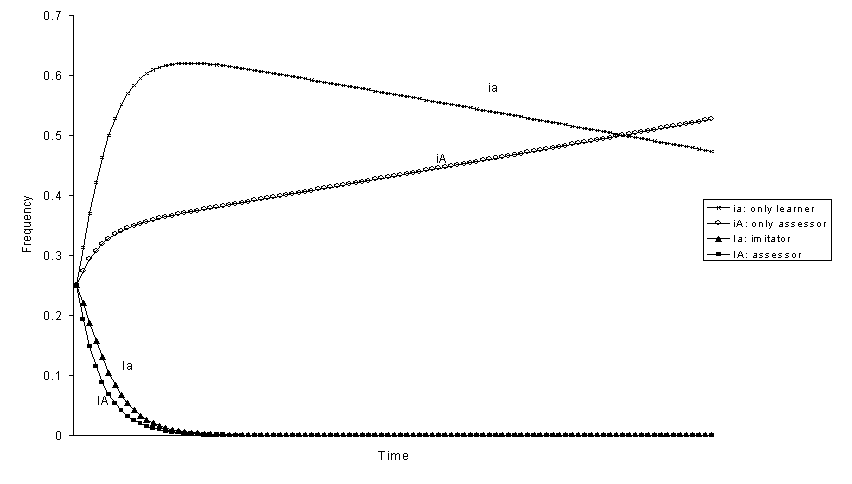
Figure 5. Evolution of genotypic frequencies with negative selection
against phenotype 1 ![]()
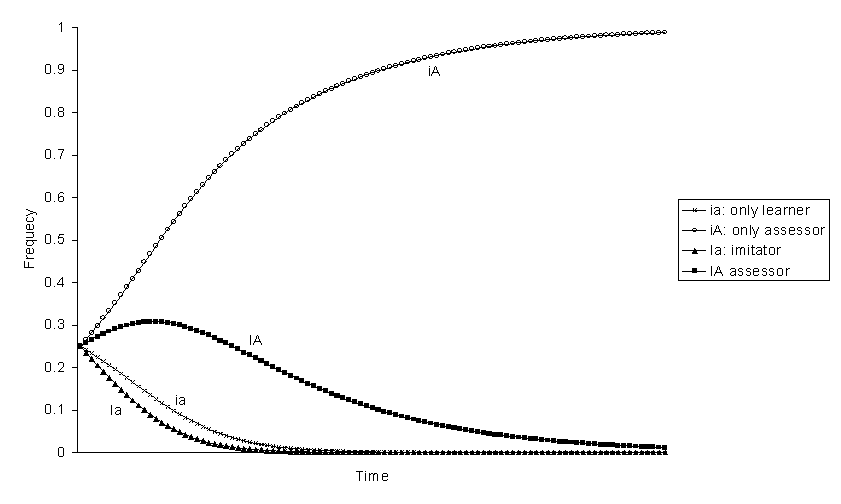
Figure 6. Evolution of genotypic frequencies with negative selection
against phenotype 1![]()
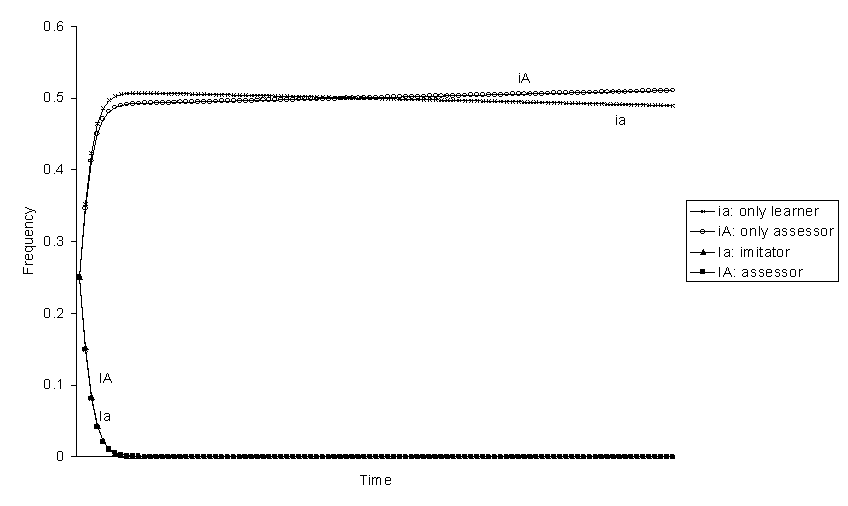
Figure 7. Evolution of genotypic frequencies with negative selection
against phenotype 1 ![]()

Figure 8. Evolution of genotypic frequencies with negative selection
against phenotype 1 ![]()
If the approval or disapproval of the behaviour exhibited by the
offspring has a cost c, assessor and only-assessor can lose their
advantage in some of the previous situations. The cost comes from the time
and energy devoted to influence the offsprings behaviour. Obviously, this
will depend on the benefit of evaluation outweighing or not the cost. However,
it is important to bear in mind that a given amount of parental investment
can be used to control simultaneously several behaviours learned by the
offspring. Thus the cost will be attenuated because it will be offset by
the benefits obtained from the transmission of information on several learned
behaviours.
The main effect of imitation in the model (a
> 0) is to increase the probability that naive individuals learn the variants
1 present in the parental generation. Imitation allows individuals to have
access to variants difficult to learn by individual learning, irrespective
of the final decision to accept or reject the behaviour. Consequently,
the effect of imitation is stronger when the behaviour is more difficult
to learn by trial and error learning -low values of h.
In our model, imitation is favoured by natural selection -fitness increases as a increases- whenever the imitated variant 1 has a positive selective value and it is at a disadvantage if the behaviour that can be imitated 1 has a negative fitness value (s < 0). The imitation will be adaptive if the parental behaviours are mostly adaptive. In a more realistic model that considers the possibility of imitating other individuals different from the parents -oblique and horizontal cultural transmission- the probability of knowing any behaviour present in the population will increase and it will approach 1 if the population is small and the number of behaviours is not too great. Therefore, almost all of the individuals will know all the variants present in the parental generation and they will have to decide which of them they accept. In this situation, the crucial aspect for adaptation is the preference of the individuals for the variants with greater fitness or, in other words, a positive correlation between b and fitness.
Our model assumes an asymmetry with respect to the imitation of both variants: variant 1 can be imitated but variant 0 cannot. In some situations this assumption is not realistic because both behaviours can be imitated and can be treated as allele-phenotypes. In these cases, we have shown that imitation is always favoured by natural selection when the parameter a is the same for both phenotypes 1 and 0 (Castro & Toro, 1995).
The model does not distinguish between individuals that really imitate
the behaviour they observe -true imitation- and individuals that manage
to develop the behaviour by indirect observation -i.e. stimulus enhancement.
Both mechanisms raise the probability that individuals will perform and
test a behaviour, but do not modify the probability of accepting it. If
stimulus enhancement is as efficient as imitation in raising the probability
of performing a behaviour, the fitness variation caused by the two mechanisms
will be equal.
The only-assessor individual can approve or disapprove of the behaviour
of their offspring. Only-assessor transmits evaluations about behaviours
he has learned and categorised previously, in a process of cultural transmission
distinct from imitation and that can be called teaching in a strict sense.
This transmission can have a cost c that decreases the fitness of
only-assessor.
The transmission of the evaluation of behaviour 1 increases or decreases
the final probability b* of
offspring preferring variant 1 instead of 0, but also increases
the phenotypic correlation between parents and offspring. Thus, only-assessor
will always be better than only-learner because he increases the frequency
of the adaptive behaviour, 1 or 0. Only if the cost c of the transmission
of evaluations is great enough, only-assessor can become worse than only-learner.
However, the ability to approve or disapprove of offsprings behaviour
is not present in non-human animals. Chimpanzees and other primates do
punish behaviour of others but only if this behaviour affects them directly
and negatively and this does not require a conceptual categorisation of
behaviour. We think that the emergence of this ability required the previous
development of the conceptual capacity to categorise learned one's own
and others' behaviour as 'positive' or 'negative'.
The assessor genotype has both abilities, to imitate and to transmit
evaluations. In the model, assessor is the best genotype when the learned
behaviour that can be imitated is adaptive (s>0). If behaviour 1
is maladaptive (s<0), genotype assessor can be the worst genotype.
The reason is that the imitation of behaviour 1 is maladaptive. However,
he can be better than imitator and even better than only-learner if the
effect of imitation is small (h close to 1) and the effect of transmission
of evaluations is great (d close to 1 and b
close to 0). Similar to what happens with only-assessor, if there is a
cost c in the transmission of evaluations the fitness of assessor
will decrease.
Regardless of how they are acquired -by trial and error, imitation or insight-, behaviours are evaluated by each one of the individuals of the population and accepted or rejected. In our opinion, social learning -stimulus enhancement or true imitation- in primates and other mammals, is not an alternative to individual learning by trial and error -older in the phylogeny- but a complementary mechanism that permits the development of quicker and with higher probability the behaviours present in the parental generation. The assessor parents can generate a more efficient system of cultural inheritance, because they transmit information on both the behaviour and the evaluation of behaviour and, thus, increase the phenotypic correlation between parents and offspring. The cultural transmission that produces assessor is however a flexible inheritance system, because it allows the propagation of innovations. If bis close to 1, a new behaviour that can be imitated through oblique or horizontal transmission will be easily propagated, because parents cannot disapprove of an unknown variant that it is not present in their status. In this way, the efficacy of vertical cultural transmission is compatible with a quick dissemination of innovation through oblique or horizontal transmission.
The existence of biases in the assessment can notably increase the efficiency of assessor. It seems reasonable to admit that individuals can transmit -approve or disapprove of- information on the value of a behaviour with more or less intensity -greater or lower d - depending on the ease or difficulty of the decision process, and on the confidence they have on the decision taken. The biased assessment could occur without any genetic differences among individuals in the value of d . We have put forward the idea that the intellectual capacity of the human species attained its extraordinary development within a framework of social life and cultural transmission of values, as a new structure capable of generating values and showing preferences between different kinds of behaviour (Castro & Toro, 1995; Castro & Toro, 1998; Castro et al., 1998). We think that the cultural transmission of values determined the propagation of memes leading to a process of cultural selection (i.e., the differential survival and reproduction of memes).
The distinction between b and d
values helps one understand why an individual can (or cannot) exhibit a
behaviour and, at the same time, he can disapprove of (or approve of) this
behaviour in other individuals. This property can be useful because it
allows the reproof of the imitation of dangerous behaviours before the
offspring is prepared to perform them. Moreover, it can produce parental
manipulation -i.e. penalising behaviour that serves the offspring's interests,
but does not serve the parents'.
Cultural transmission has attained an extraordinary development
in our species and is determinant of its own evolution. Human cultural
learning created a species-unique process of cumulative cultural evolution.
Boyd & Richerson (1996) argue that humans and other primates both engage
in the same kinds of social and imitative learning, but there is a quantitative
difference in social learning skills that leads to a qualitative difference
in the cultural traditions of chimpanzees and humans. On the contrary,
Tomasello (1999) suggests that human beings evolved a new form of social
cognition, the ability to "identify" with conspecifics, which led to an
understanding of them as intentional and mental beings like themselves,
which enabled a new process of cumulative cultural evolution. This process
depends on innovation, imitative learning, and perhaps active instruction
on the part of adults, and cannot be brought about by means of weaker forms
of social learning. We maintain that the transition between the chimpanzees'
rudimentary cultural transmission -a mixture of stimulus enhancement and
true imitation- and the human cultural transmission -based on true imitation
and teaching, defined as the transmission of information on the evaluation
of the learned behaviour - required the development of the capacity to
approve or disapprove of offsprings behaviour and previously of the conceptual
capacity of categorise learned behaviour. Or, in other words, that some
of our ancestors became assessor. The approval or disapproval of offsprings
behaviour is a new mode of cultural transmission that increases the accuracy
of the process of meme imitation, or in Blackmore (1999) words, the process
of 'copy-the-product'. Thus, cultural transmission becomes a system of
inheritance and cumulative cultural transmission become possible.
In summary, the success of assessor is due to his capacity to increase the phenotypic correlation between parents and offspring, amplifying the effect of natural selection on learned behaviours -favouring an increase in the adaptive ones and disfavouring the maladaptive ones. Assessor parents can do one important thing that imitators cannot: they can help the offspring to quickly and firmly reject maladaptive behaviour with negative evaluation or adaptive behaviours that are maladaptive at their age. Moreover, assessors can also facilitate the quick propagation of innovations and of the adaptive behaviours when they acquire a positive bias in the cultural transmission of evaluations.
Let us consider a population monomorphic for the most complex genotype
IA. If f00, f01 and f11
are the phenotypic frequencies in generation t, the values g00,
g01
and g11 in generation t+1 will be

At the equilibrium the frequencies will remain unchanged
![]() .
.
After some algebraic manipulation it can be shown that these equilibrium
values will be,

The equilibrium frequencies for the less complex genotypes can be easily calculated from the above equations assuming d =0 (genotype Ia), a =0 (genotype iA) and a =0 , b =0 (genotype ia).
Thus, for the genotypes ia and iA the equilibrium values will be

and for the genotype Ia
This work has been developed within the R+D Project: Estudio
interdisciplinar de rasgos funcionales del grado humano (BSO2000-116-C04).
The work has also been facilitated by concession to L. Castro of a licencia
por estudios of the local Government of Madrid.
Blackmore, S., 1999. The Meme Machine, Oxford, Oxford University Press.
Boyd, R., & Richerson, P. J., 1985. Culture and the Evolutionary Process, Chicago, The University of Chicago Press.
Boyd, R., & Richerson, P. J., 1995. Why does culture increase human adaptability? Ethology and Sociobiology, 16,125-143.
Boyd, R., & Richerson, P. J., 1996. Why culture is common but cultural evolution is rare? Proceedings of the British Academy, 88, 77-93.
Byrne, R., & Russon, A. E., 1998. Learning by imitation: A hierarchical approach. Behav. Brain Sci., 21, 667-721.
Castro, L., 1992 .Capacidad ética, transmisión cultural y evolución humana. Arbor, 564, 81-92.
Castro, L., & Toro, M. A., 1995. Human Evolution and the Capacity to Categorise. Journal of Social and Evolutionary Systems, 18, 55-66.
Castro, L., & Toro, M. A., 1998. The long and winding road to the ethical capacity. History and Philosophy of the Life Sciences, 20, 77-92.
Castro, L., Serrano, J.M., & Toro, M. A., 1998. Conceptual capacity to categorize and the evolution of altruism. Journal of Theoretical Biology, 192, 561-565.
Custance, D., Whiten, A., & Fredman, T., 1999. Social learning of an artificial fruit task in capuchin monkeys (Cebus apella). J. Comp. Psychol., 113, 13-23.
Feldman, M. W., & Cavalli-Sforza, L. L., 1984. Cultural and biological evolutionary processes: Gene-culture disequilibrium. Proc. Natl. Acad. Sci. USA, 81, 1604-1607.
Feldman, M. W., Aoki, K., & Kumm, J., 1996. Individual versus social learning: Evolutionary analysis in a fluctuating environment. Anthropological Science, 104, 209-232.
Galef, B.G., 1988. Imitation in animals: History, definitions, and interpretations from the psychological laboratory. In Social Learning: Psychological and Biological Approaches. T. Zentall & B.G. Galef (Eds.) Hillsdale, NJ: Lawrence Erlbaum Assoc.
Galef, B.G., 1992. The question of animal culture. Human Nature, 3, 157-178.
Henrich, J., and Boyd, R., 1998. The evolution of conformist transmission and the emergence of between group-differences. Evolution and Human Behavior, 19, 215-241.
Heyes C.M., & Galef, B.G., JR. (Eds.), 1996. Social Learning in Animals: The Roots of Culture, New York, Academic Press.
Kendal, J. & Laland, K., 2000. Mathematical models for memetics. Journal of Memetics - Evolutionary Models of Information Transmission, 4 (1), http://cfpm.org/jom-emit/2000/vol4/kendal_jr&laland_kn.html.
Maynard Smith, J., & Warren, N. 1982 "Models of cultural and genetic change". Evolution, 36: 620-627.
Rogoff, B., 1990. Apprenticeship in thinking: Cognitive development in social context, Oxford, Oxford University Press.
Tomasello, M., 1990. Cultural transmission in the tool use and communicatory signaling of chimpanzees. In "Language" and Intelligence in Monkeys and Apes. Comparative and Developmental Perspectives. S.T. Parker & R.K. Gibson (Eds.) Cambridge, Cambridge University Press.
Tomasello, M., 1996. Do apes ape? In Social Learning in Animals: The Roots of Culture. C.M. Heyes & B.G. Galef (Eds.) New York: Academic Press.
Tomasello, M., 1999. The Cultural Origins of Human Cognition, Cambridge, Mass, Harvard University Press.
Tomasello, M., Kruger, A. C., & Ratner, H.H., 1993. Cultural Learning. Behav. Brain Sci., 16, 495-552.
Visalberghi, E., & Fragaszy, D. M., 1990. Do monkeys ape?. In "Language" and Intelligence in Monkeys and Apes. Comparative and Developmental Perspectives. S.T.
Whiten, A., 1998. Imitation of the sequential structure of actions by chimpanzees (Pan troglodytes). J. Comp. Psychol., 13, 270-281.
Whiten, A., Goodall, J., McGrew, W.C., Nishida, T., Reynolds, V., Sugiyama, Y., Tutin, C.E.G., Wrangham, R.W., & Boesch, C., 1999. Cultures in chimpanzees. Nature, 399, 682-685.
Whiten, A., & Ham, R., 1992. On the nature and evolution of imitation
in the animal kingdom: reappraisal of a century of research. Adv. Study
Behav., 21, 239-283.