 Wilkins,
J. S., 1998; What's in a Meme? Reflections from the
perspective of the history and philosophy of evolutionary biology.
Journal of Memetics - Evolutionary Models of Information
Transmission, 2. http://cfpm.org/jom-emit/1998/vol2/wilkins_js.html
Wilkins,
J. S., 1998; What's in a Meme? Reflections from the
perspective of the history and philosophy of evolutionary biology.
Journal of Memetics - Evolutionary Models of Information
Transmission, 2. http://cfpm.org/jom-emit/1998/vol2/wilkins_js.html
What's in a Meme?
Reflections from the perspective of the history and philosophy of evolutionary
biology
John
S. Wilkins
3 Peel Grove, Mt. Martha 3934, Australia.
Contents
- 1 - Introduction
- 2 - The background to the problem
- 3 - The Hull-Dawkins Distinction
and the evolutionary gene
- Sidebar: interactors and replicators
- Sidebar: phenotypes, phemotypes and classification
- 4 - The lineages and ecologies
of culture
- 5 - Memetic individuals
- 6 - Conclusion and prospectus
- Acknowledgements
- Glossary
- Notes
- References
Abstract
This paper is intended as a focal article to
raise philosophical issues about the nature of memes and memetic theory.
To bring consistency to memetic analysis, researchers need to understand
and agree upon the theoretical role of memes and the generalized model
of evolution in which it occurs as a theoretical term. To help this, I
have traced the source of Dawkins' conception of memes from GC Williams'
evolutionary gene and through to the Hull-Dawkins Distinction between
replicators and interactors and Hull's notion of lineages
and the idea of an individual in biology. The complexity of biological
modes of evolution suggests that conceptualizing memes as disease pathogens
is not an alternative to evolutionary models of memetic development. I
argue for a close and strict analogy between biology and memetics. I introduce
the idea of a memetic individual or profile to clarify the
ontology of memes and their ecologies. Some promising methods from biology
and other disciplines such as Hamming Distance and Wagner groundplan divergence
methods are suggested. A glossary of mainly biological technical terms
used and introduced neologisms is included.
Keywords: Meme, evolution, gene, replicator, interactor, individual,
natural selection, term, mnemone, pheme, deme, ecology, epidemiology, mind
virus, Lamarckism, Dawkins, Hull, Campbell, Williams
JULIET: 'Tis but thy name that is my enemy;
Thou art thyself, though not a Montague.
What's Montague? it is nor hand, nor foot,
Nor arm, nor face, nor any other part
Belonging to a man. O, be some other name!
What's in a name? that which we call a rose
By any other name would smell as sweet;
So Romeo would, were he not Romeo call'd,
Retain that dear perfection which he owes
Without that title. Romeo, doff thy name,
And for that name which is no part of thee
Take all myself.
(William Shakespeare, Romeo and Juliet, Act 2, Scene 2)
1 Introduction
The fundamental theoretical concept of memetics is the meme
- the unit of cultural evolution and selection. The term is unclear in
its meaning and what it denotes, and the application of evolution to culture
is often based on a partial or even mistaken notion of the general structure
of evolutionary explanation. In this paper I give some historical background
to the origin and function of the term "meme" from the debates
in evolutionary theory out of which it sprang. I argue that there is a
close analogy between the entities and processes of biology and culture,
although the parameters that describe each domain often differ in their
rate and frequency, and perhaps in the dynamics of the evolutionary process
they undergo. It is as crucial to understand what the units of evolution
in culture are as it is in biology to be able to apply an evolutionary
explanation, and for this reason we must be quite clear about what a cultural
individual is. We can make better sense of these issues if we look first
at the analogous problems of what a biological individual is, especially
in the context of what the units of selection are.
Problems that bedevil memetics include classifying cultural entities
and reconstructing historical developments, and these also have their analogues
in biology. Resolving them depends on determining the level at which selection
occurs, for it is selection that defines what is functionally important
in evolution, not the contrary. The conceptual tool of most help to us
here is the distinction between replicators
and interactors. There are
three common approaches in memetic writings - most commonly writers picture
meme evolution as the spread of disease pathogens, others restrict selectionist
accounts to some part of culture that is amenable to this form of explanation
such as science, and I argue for a third option. We must more broadly understand
the variety of biological evolutionary phenomena, so that we can see the
similar patterns in cultural change. In order to assist in this, I introduce
the notion of a memetic
individual, and discuss the relationship this entity has with the biological
organism in which it resides. In the conclusion and prospectus, I suggest
some analytical techniques of biology, particularly methods of taxonomy
and genetics, which these distinctions permit for memetic research.
I hope both that biologically literate readers will understand my simplifications
of biology and that those not familiar with biological terminology its
necessity. I append a glossary of various technical
terms introduced in this paper and those that are likely to be unfamiliar
to nonbiologists.
2 The background
to the problem
The increase of the conceptual clarity of a theory through careful
clarifications and specifications is, as William Whewell observed more
than a century ago, one of the most important ways in which science progresses.
He called this process "the explication of conceptions" and showed
how a number of theories, in the course of their temporal careers, had
become increasingly precise - largely as a result of the critics of such
theories emphasizing their conceptual unclarities. Many important scientific
revolutions ... have depended largely on the recognition, and subsequent
reduction, of the terminological ambiguity of theories. (Laudan
1977:50)
Certain terms and notions in both the sciences and humanities are fated
to be misunderstood, either because they are first vaguely formulated,
or because they are so evocative they generate such immense enthusiasm
and are applied to almost everything, coming to mean almost nothing. A
classic example is the term of Thomas Kuhn's (1962):
paradigm. Originally intended by Kuhn to apply to what changed radically
in a scientific revolution, it came to be applied to perceptual and conceptual
changes in cases of individual, social, literary, political, economic and
even consumer choice. When a term of philosophy of science is used to advertise
a new car design, you know it has lost any definite meaning. Eventually
its author abandoned it under criticism in favour of notions and terms
that were more specific, but "paradigm" is now ensconced in popular
parlance, surviving both author and intended theoretical usage. The difficulty
now with the term for a specialist in the philosophy and history of science
is that calling a theoretical change a "paradigm shift" has become
little more than a metaphor. It describes only an impression and implies
only a subjective assessment.
The basic and central notion of memetics is, of course, denoted by the
term meme, Richard
Dawkins' (1977) term for what is transmitted in culture that is analogous
to the biological gene. "Meme" is in danger of suffering the
same fate as "paradigm". It is used to denote, variously, neural
structures, cultural artefacts, practices, economic systems, religions,
concepts, phenotypic traits, self-awareness, and epigenetic predispositions.
Memes are thought by some to control behaviour, by others to be acquired
through a choice or act of will. The term gets applied to all levels of
social and cultural structure, from minimal semantic entities like phonemes,
through more molecular entities like phrases and snatches of music, to
entire traditions and world views. In this blooming buzzing confusion,
the usefulness of memes as a category is being lost or degraded.
It wouldn't be the first time this sort of confusion has wrecked an
emerging discipline - gene, the very
term on which "meme" is modelled, has a history of shifting definition
and debates in which antagonists argued past each other for more than sixty
years. It took the modern evolutionary synthesis and the discovery of the
structure of DNA to resolve it fully, and still, "gene" covers
a range of theoretical phenomena requiring the more exact terminology of
codons, cistrons, introns, exons, operons, regulators and base
pairs. Even today, biologists use this term differently and inconsistently
- what molecular biologists usually mean by it differs greatly from its
meaning in population genetics.
A theoretical term is usually generated to denote a causal nexus in
the model the theory describes.
On at least one recent account (Suppe 1989, van
Fraassen 1980) a scientific theory is an attempt to either isolate
or idealize a system - usually a physical system - in such a way that its
dynamics can be reduced to a manageable number of variables (each of which
is usually represented by a theoretical term) related by a mathematical
description, so that the model generates a restricted number of likely
outcome states. It is often neutral with respect to many of the attributes
of the entities and processes it covers: Newton's theory of falling bodies
does not mention their colour, for example; the equations of flow dynamics
cover water, hydrocarbon liquids, and gases; and information theory covers
transmission of bits by electromagnetic radiation no matter whether the
medium is electrical, photic or magnetic.
One such generalized scientific theory is the theory of adaptation and
evolution by natural selection. Natural selection is a process that occurs
over a range of physical substrate entities: viruses, single celled clonal
organisms, multicellular plants, animals and fungi, and so forth. Certain
variables need to be adjusted to deal with these very different phenomena
- life cycle rates, ecological behaviour, reproduction modes and so forth
- but the virtue of the theory as a model is that it is highly generalizable
and widely applicable. Recently, for example, natural selection models
have been applied through the use of computers to a range of real world
problems, and have been developed into a class of formal algorithms called
Genetic Algorithms (cf. Holland 1995). Problems
to which Genetic Algorithms have been applied include political and social
problems, through to hard engineering of complex systems like engines and
flow dynamics.
To understand the meaning of meme, we need to understand the
history of some debates in evolutionary biology, and the ways in which
evolution and culture have been thought to be related. Even before Darwin
published the Origin in 1859, evolutionary
reconceptualisations of human society and culture were common. The Lamarckian
evolutionary tradition through Geoffroy, Grant and Chambers had drawn radical
social conclusions based on their view of evolution as progressivist, universal
and hierarchical (Desmond 1989). Not long before
the Origin, Herbert Spencer developed his form of Hobbesian rugged
individualism based on a Malthusian struggle for existence, ideas that
found a parallel in Darwin's natural history work. But of interest to a
memeticist is the application of natural selection models to ideas, by
writers like Alexander Bain, Charles S Peirce, William James, Ernst Mach,
and John Dewey, very soon after Darwinism was first promulgated through
European intellectual society. In many ways, Dewey is one of the first
true memeticists, declaring that we didn't so much solve problems as recover
from them, appropriating both the Darwinian evolutionary metaphor and the
epidemiological metaphor so popular today (Dewey 1909).
Over the years, various others have used natural selection as a metaphor,
or as an analogy (the distinction is significant
), of conceptual and social change, including Medawar, Popper, Toulmin,
and even some leading Darwinians (cf. Cziko 1995
for a review).
Eugenics was a constant source of confusion and debate in the early
synthesis, with Fisher and the "Oxford School" along with leading
geneticists like Muller proposing that the primary role of natural selection
was to trim away deleterious alleles and that the best way to organize
society was to permit that process to act without impediment. Dobzhansky
and others, largely in North America, argued instead that the role of natural
selection was to maintain a pool of variety in populations, and that equality
of opportunity would permit whatever alleles had relative advantage to
be expressed (Beatty 1987). This was more significant
than a clash of political or moral doctrines. Eugenicists (Darwinian eugenics
was usually positive - encourage the fitter - than negative - cull the
less fit - as in Spencerian social "Darwinism") held that society,
or at any rate "civilized" society, acted to lessen the effects
of natural selection, while those who like Dobzhansky opposed eugenics
tended to see society as either neutral or ancillary to natural selection
in biology. The very role of natural selection itself was the subject of
intense debate, culminating during the final years of the synthesis and
the early 50s. It centred on an extended debate between Fisherians, who
saw it as a universal and often all-sufficient mechanism of evolutionary
change, and those who like Dobzhansky followed Sewall Wright in thinking
that in small populations (demes) random drift and sampling error played
a greater or lesser role in the evolution of traits. Even today this is
still an issue. Dobzhansky, who through his early education in the tradition
of Russian Darwinism which emphasized co-operation and began with Kropotkin,
was well placed to argue for Sewall Wright's notions, and by the end of
the 1950s, some admixture of drift and selection was accepted by nearly
every evolutionary biologist.
The point of this potted history is to show the context in which the
meme-analogue "gene" arose and was debated, and to set the scene
for the conditions in which memes were developed as theoretical entities;
this will go to the question of the nature of memes and their theoretical
role in explanation. Throughout the period of the development of Darwinism,
it was agreed that there had to be hereditary factors of some kind because
natural selection can only act if the traits of organisms that are selected
are transmitted to future generations in a process of differential reproductive
success based on what we now call differential ecological success. But
it took a long time for this to be clarified. Evolutionists often spoke
of new traits being "for the good of the species" even though
Fisherian genetics modelled fitness as an individual property of
heritable traits in a population. Indeed, Fisherian genetics treated genes
in such an unlinked atomistic way that later writers like Mayr spoke derisively
of "bean-bag" genetics, which treated genes as if any single
one of them could be separated out from the others and inspected. Although
Mendelian genetics had already realized that genes were often linked in
their effects, and that many genes affected each trait ( epistasis
) and some genes could affect many traits ( pleiotropy
), it was unclear what natural selection acted on - individual genes
or the traits they affected. In his 1937, Dobzhansky adopted Darwin's term
"co-adaptation" to denote parcels of genes that were selected
as a unit because they had evolved together to work well as a team, but
there was a real epistemological difficulty in determining what genes selection
actually selected.
In 1962, the modern "units of selection" debate was inadvertently
kicked off by a naturalist, Wynne-Edwards, who explained the regulation
by some gulls of clutch size (the number of eggs laid) during lean times
as an adaptation that could only be for the benefit of the group, because
individual fitness had to be lower if fewer progeny were produced. Wynne-Edwards
used the notion of "group selection" which Darwin had used in
1872 to explain the existence of human moral behaviours: moral groups did
better than immoral groups, and so their genes would be better represented
later on than those of the others (to reinterpret Darwin's formulation
in modern terms). According to Wynne-Edwards, populations that regulated
clutch size survived ecological catastrophes better than those that didn't.
It's an intuitively seductive idea, but this sort of reasoning was thought
to be woolly headed and wrong by those evolutionary theorists raised in
the more reductive panselectionist tradition. David Lack argued against
the biological specifics of Wynne-Edwards' thesis, but a pivotal outcome
of this book was that it spurred George Williams to publish, in 1966, Adaptation
and Natural Selection.
Williams took an operationally reductive approach; that is, he refused
to use selectionist explanations at any level higher in the biological
hierarchy than was necessary. It is onerous, he thought, to explain group
level phenomena in sui generis terms when explanation in terms of
the components of the group and their properties will suffice. For
selection of a group to occur (and contrary to popular reportage, Williams
did not rule out the in-principle possibility of group selection, even
identifying a case study), two criteria would need to be satisfied. One,
the properties that were favourably selected would need to be the properties
of the group, not the individuals that comprised it. Williams illustrated
the difference by distinguishing between a group of fleet deer (which as
a group may move slowly, though all its parts are fast moving) and a fleet
group of deer (which may move quickly, as groups go, although its parts
are slower than parts of slower groups). Fleet deer are individuals with
properties of fleetness. Fleet groups of deer are groups with properties
of fleetness. The second criterion that must be satisfied is that group
properties had to be heritable: copies must be made if selection is to
operate. Therein lay the rub for Williams: groups typically do not reproduce.
But Williams went even further: neither are individual organisms usually
copied. You are not a clone of your same sex parent. You may have your
father's eyes, your mother's hair colour and wave, and be intermediate
between them in your genetic predisposition to height. Mendelian factors
are the basic unit of evolution, according to Williams, and not all of
them, either. "I use the term `gene' to mean that which separates
and recombines with appreciable frequency", he said (1966:
20) and he further defined an evolutionary
gene (that is, one that is selectively important) as
"...any inherited information for which there is a favorable
or unfavorable selection bias equal to several or many times its rate of
endogenous change" (1966: 25).
In set theoretic terms: Mendelian factors (M) are a proper subset of
heritable separating and recombining units of information (G), and include
as a subset selectively biased - that is, evolutionary (E) - factors:  .
A Venn diagram of this relationship shows how he appeared to view it:
.
A Venn diagram of this relationship shows how he appeared to view it:

Ten years later, Richard Dawkins (1977) introduced
the "selfish gene" metaphor based on Williams' "evolutionary
gene". But the way he did so obscured the point Williams was making
(not deliberately, for Dawkins had other fish to fry) - not all genes are
evolutionary genes. Given that only genes can be evolutionarily relevant,
what portion of those genes actually are? Williams is a bit of a
panselectionist and he
tends here to brush past questions of random
drift and founder effects.
His answer to the question is that a gene is evolutionary just insofar
as it is subject to selection that exceeds mutation. This is a definition,
not a discovery. If it's favourably or unfavourably selected and heritable,
it's an evolutionary gene. We are now on the threshold of memes. Williams
himself realized this, noting that gene was a cybernetic abstraction, and
later describing the notion of a "codical domain" of evolution,
wherein the transmitted structure, no matter what its physical substrate,
is a codex (Williams 1992). In other words,
it's a message that is transmitted, and which is subjected to the same
theoretical models as any other kind of message transmission, i.e., Shannon-Weaver
information constraints. This means that genetic replication is one instance
of a class of phenomena in Message World - and memes, like genes, are another.
Dawkins' original introduction of the term "meme" in The
Selfish Gene mentioned in passing snatches of tunes, crazes and
fads, but the paradigmatic example he gave, no doubt due to his personal
experience of it, was a scientific notion passed from scientist to scientist.
I shall return to this point later, but some properties of scientific notions
are of immediate interest. Typically, scientific ideas are either evocative
metaphors, like de Candolle's "struggle for existence" that inspired
Darwin, or more or less formal models. It is the latter that concerns us
here, for when metaphors reach the end of their evocation, they must be
turned into formal models anyway in order to be tested against quantifiable
phenomena. A formal model like Boltzmann's thermodynamic entropy is a far
cry from Heraclitus' notion of flux, and it does a great deal more conceptual
work. The significance of Dawkins' example is that one can, to a relative
degree of exactness, determine whether and how far a part or whole of a
model has spread to another scientist or textbook, or whatever one takes
to be the cultural equivalent of the phenotype - for reasons I hope become
clear, I shall refer to this as the phemotype,
and the total distribution of co-adapted memetically constituted traits
within a lineage the phemorph
of that lineage. The neologisms are strained, barbarous and ugly, but I
hope they will add some clarity to what is being discussed. In short, scientific
examples can be quantified both in terms of their frequency in a lineage
of scientists, and their relative rates of increase or decrease. This susceptibility
to analysis is essential for modelling change in terms of natural selection,
and evolution in general.
3 The Hull-Dawkins
Distinction and the evolutionary gene
Do I contradict myself?
Very well, I contradict myself,
(I am large, I contain multitudes.)
Walt Whitman, Song of the Open Road
To apply evolutionary models outside biology, you need to know what
evolution as such is, no matter what the domain it is occurring in. There
have been many attempts within biology to generalize evolutionary modelling,
for it is a point worth remembering that even biological evolution is not
all of a kind. Evolution occurs over single and multicellular organisms,
over animals and plants and fungi, and over various kinds of bacteria and
viruses. It applies to sexual, clonal and parthenogenic
reproduction, to heredity using DNA, RNA and even cytological structure.
Generalist species (eurytopes) and specialist species (stenotopes)
both evolve in widespread and continuous ecologies, and in localized and
isolated ecologies, and so on. Evolution is not simple in biology, and
so biologists have worked to make it as general a model as possible to
cover all cases. There is clearly some common thread in all these phenomena,
and most find some or all of the explanation in natural selection.
As Weismann's and Wallace's heirs perceive natural selection, it is the
differential success of the genotype due to the ecological success of the
phenotype it creates. The causal arrow is essentially one way: genes cause
phenes but not the reverse. As Cziko
(1995), following Donald Campbell, puts it, genes are selected but
not instructed. Despite all the progress since Weismann on the nature of
biological heredity, his Central
Dogma has stood almost without modification.
It is therefore important to determine whether there is an analogue in
culture for these central distinctions of genotype and phenotype and whether
memes are ever instructed, or only ever selected, as genes are. And if
memes are instructed, does this "Lamarckism"
of culture obviate the need for a memetic analysis? Although it's worth
remembering that Darwin was a Lamarckian on heredity, that is, he accepted
that the units of heredity ("pangenes") are instructed, this
does not really help us answer the question, for we already have non-evolutionary
Lamarckian models of culture and the justification for taking an evolutionary
perspective rests on the efficacy of Darwinian, perhaps Weismannian, models.
We need to understand the ontology of a Darwinian process to come to any
resolution.
Dawkins (1982) distinguished between genes
as replicators - things that
are duplicated with a high degree of fidelity - and bodies of organisms
as the vehicles of genes. Vehicles reproduce, but only genes replicate.
Some writers, including David Hull (1980, 1987,
1988a, 1988b, 1988c)
took exception to Dawkins' characterization of bodies as mere vehicles
of genes - they were, in his opinion, much more than passive and controlled
robots. Hull had earlier
(1974) attacked explanatory reductionism
in genetics, arguing that reducing evolutionarily significant traits to
genes was unsatisfactory both in terms of a failure of isomorphic relations
(mapping) and also of underdetermination - things happen at metagenic levels
that are not, and never could be, the results of the sum of the properties
of the genes. He therefore
rejected Dawkins' passive notion of bodies and the phenotype, and substituted
instead the more ecologically and economically active notion of an interactor.
Hull's general view of evolution is of a cycle of replicators coding for
interactive traits, which through their interactive success acquire (or
fail to acquire) the resources needed for further replication. On Hull's
account, ecology makes a strong comeback, and the reproduction of organisms
is a necessarily interactive, rather than a replicative,
process. This cycle generates lineages
of descent, which exhibit themselves in biology as species in the short
term and as phylogenies and other taxonomic classes in the longer term.
Hull and Ghiselin, as part of their individuality thesis that
species are historical individuals, that happen to be made up of many organisms,
have also addressed the general problem of what constitutes an individual
in biology and evolution in general (Hull 1987,
1988c, 1992, Ghiselin
1997). This also raises its head in memetics, and has not so far been
addressed. Their view is that a biological individual, so far from being
intuitively obvious (as it seems to be if we only consider obligately
sexual vertebrates) is in fact a very fluid category, and their solution
is that an individual is defined by its functional role in evolution,
that is, insofar as it is a unit of selection (this applies to gene sequences
as well as to organisms). So, the resultant evolutionary ontology is that
replicators (replicanda in Ghiselin's terminology - structures that are
replicated, Ghiselin 1987) generate interactors,
which are the evolutionary individuals that are subjected to selection
and whose economic success or failure biases the regeneration of the replicanda.
Plotkin (1994) characterized generalized selective
processes as generate-test-regenerate cycles, and this helps to
clarify the matter (figure 1). An individual is formed
at the level of testing, which feeds back into the generation stage. An
inclusive hierarchy of individuals can be formed if generation to regeneration
cycles occur at higher levels.
In biological terms, this accounts for why colonies of organisms like eusocial
insects behave as individuals with respect to selection, and also why clonal
lineages (genets) behave evolutionarily
as individuals even if they are spatially distinct (ramets).
|
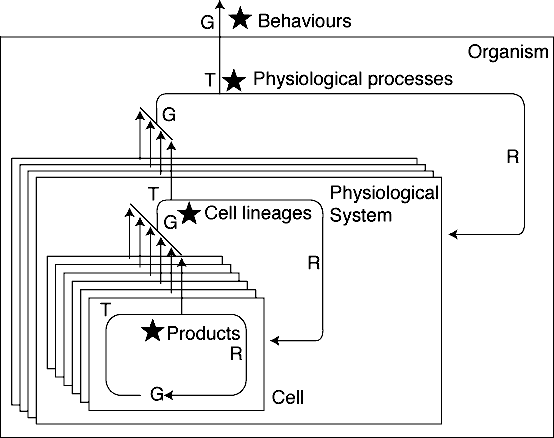
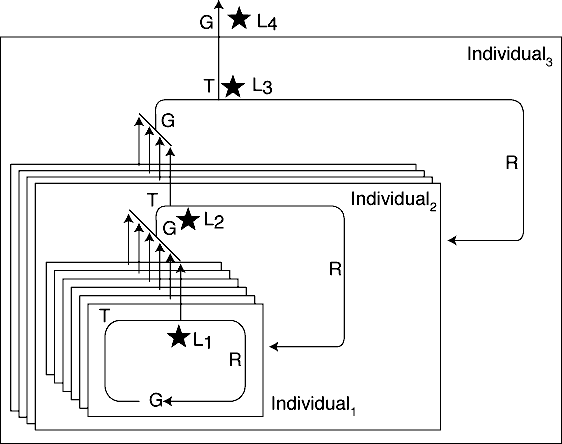 Figure 1. A generalized hierarchy of g-t-r
(generate-test-regenerate) cycles in an evolutionary process. Star = Replicators
relative to testing/selection bias at that level or higher
Figure 1. A generalized hierarchy of g-t-r
(generate-test-regenerate) cycles in an evolutionary process. Star = Replicators
relative to testing/selection bias at that level or higher
|
The Hull-Dawkins Distinction, as it came to be called, gained almost
immediate and bipartisan support across the selectionist debate - both
Eldredge (1989) and Williams
(1992) accepting that replicators and interactors are the active and
general entities in Darwinian evolution (interestingly, both made the now
almost obligatory passing comments about these terms also applying to culture.)
Dawkins' "memes" (like Campbell's mnemones,
1974, 1988) are
cultural replicators, which, if they are to function in cultural
evolution analogously to genes, must be transmitted with fidelity, and
must cause some interactive traits that in turn will cause a differential
replication of the memes. This also helps us with the question whether
memes are instructed or selected; that is, whether they arise in response
to environmental needs (are "learnt") or are generated randomly
with respect to the prevailing social ecology ("random trial and error").
Unexpressed memes (memes not "visible" to the environment through
their products) cannot be selected, and so the likelihood of them being
prescient or anticipatory is reduced, since we would need to have already
selected some memes as likely candidates for success in order to predict
which ones will work in practice.
Before a meme can be assessed as "likely to succeed", it must
already have passed some tests. We therefore get a regress - any "instruction"
of a meme is either a case of transmission followed by selection, or it
is a case of transmission of an already selectively tested meme.
The Central Dogma remains unshaken for memes, even if some mechanisms of
instruction are shown to occur, for even learning is a selection process
(Cziko 1995, chapters 11-12) at some level.
|
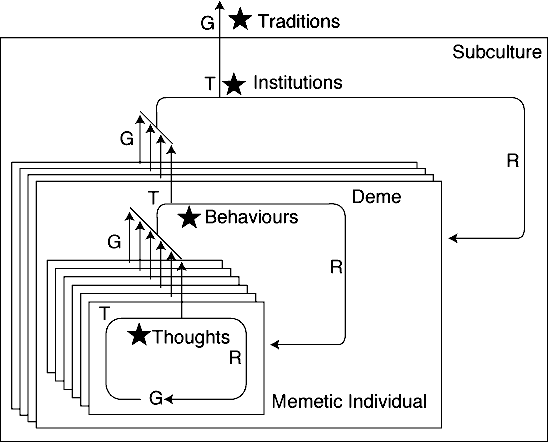 Figure 2. Memes arise as effects of a g-t-r
cycle. Star = Meme relative to testing/selection bias at that level or
higher
Figure 2. Memes arise as effects of a g-t-r
cycle. Star = Meme relative to testing/selection bias at that level or
higher
|
Some may object that memes must be stored in neural patterns
in order to be considered memes, and that replication through texts, electronic
media, and performances, etc., are only secondarily memetic if they affect
and are stored in a central nervous system. In figure 2,
it can be seen that some selectively biased cultural phenomena can exist
as the emergent properties of social systems larger than the individual,
and persist without being replicated at time frames far longer than an
individual life span. Methodological
individualism, as it is called, rejects this in favour of a more parsimonious
view, just as genic selectionists like Dawkins reject any form of group
selection (Williams' views notwithstanding). But on the Ghiselin-Hull view
of individuality in selection, group selection is not only a possibility
but an established reality, for viewed appropriately, any organism is a
group of related lineages (Buss 1987) and any gene
sequence a related group of molecules, and so on. Consider an historical
event like the Thirty Years War or an institution like a religion. Political
and religious patterns of behaviour in that war need not be stored as neural
patterns, and each of the memes of a religion need not reside in at least
one brain, for some of these memes are emergent properties of the entire
system of acting individuals, and of which they may indeed all be entirely
unaware. Were Protestants
aware of the linkage between capitalism and their theology during the rise
of modern capitalist economies, for example? If so, why did it take a sociologist
to point it out explicitly? You pays your money and you takes your choice,
for the methodological individualist debate has a long history of controversy
in sociological disciplines that continues till this day. However, the
view I am advocating here is neither individualist, nor holist, but a view
known as " emergentism
" (Nagel 1961): the doctrine that the properties of a collective whole
arise from the relationships between the properties of the components.
Simply understanding the componential properties, without understanding
the connections between them does not enable us to model the higher level
thing they comprise.
So, what are phenotypic cultural traits, the phemotypes as I
have called them? One answer, and the right one I think, has been given
variously by Campbell (1960, 1974,
1987, 1988), Hull
(1988c), Toulmin (1972) and Plotkin
(1994), to name a few, is that selection acts on behaviour, or, in
the language of the logician, on interpretation of the information contained
in a meme. Just as a gene must be expressed in order to be selectively
biased, a meme must also find expression in some way. While we often use
a verbal shorthand when we say that a gene is selected at a certain coefficient,
the biologist should never forget that genes are just the starting point
of a biochemical process which undergoes selection at a range of hierarchical
levels, from the processes of transcription in the cell to the processes
of ecological interaction in an ecosystem (figure 3).
Figure 3. Individuals and replicators in a biological
hierarchy
Sidebar: interactors
and replicators
The terms interactor and
replicator, being substantive
nouns, give the impression that there are natural kinds of things
that interact and replicate. Ghiselin (1987,
1997: 147) has argued that a better term for
the Hull-Dawkins Distinction would have been between replicanda
and interactors, that is, between those things that get replicated
and those things that are economically biased for a range of reasons. When
we understand that replication is the passing on of a message, it doesn't
really matter what the medium, or substrate, of the message is, and the
notion that there has to be some privileged replicator, like nucleotides
or neural columns, becomes unnecessary. It is less obvious that interactors
are also a functionally-defined class, but if anything interacts (and correlates
closely with replicators) then it is an interactor. It could be the organism,
but it could be some subsystem of an organism like a class of immunological
peptide, or the immune system itself, or a visual system, or it could even
be a group of organisms like a hive. There are no privileged interactors
or replicators, in biology or culture. We would do well to remember
Mill's caution:
"The tendency has always been strong to believe that whatever
received a name must be an entity or being, having an independent existence
of its own. And if no real entity answering to the name could be found,
men did not for that reason suppose that none existed, but that it was
something peculiarly abstruse and mysterious."
(Cited in Gould 1981: 320)
This modifies Hull's notion of interactor somewhat, for Hull defines
them as things, that is, as entities. Plotkin, on the other hand, resists
the "entification" of interactors (personal communication). I
do not think that Hull's own analysis requires that interactors be a unique
class of entity, such as an organism, just as long as it is an individual
(Hull 1992) with interactive (ecological, or economic)
properties. Individuals are entities, but there is no need to restrict
individuality to one level of entitivity, so long as the relevant interactive
properties are properties of that entity and not of the system of which
it is a component. However, interactors are entities even if they are not
a specific kind of entity.
|
Similarly, memes are selected at many levels through their expression
in behaviour, including verbal, practical, instrumental and intellectual,
and this behaviour need not be the behaviour of individuals; it can be
the behaviour of languages, institutions, societies and even traditions.
Consider Campbell's (1974) hierarchy of selectionist
knowledge processes:
1. Genetic adaptation
2. Nonmnemonic problem solving
3. Vicarious locomotor devices
4. Instinct
5. Habit
6. Visually supported thought
7. Mnemonically supported thought; Observational learning and imitation
8. Socially vicarious exploration
9. Language
10. Cultural cumulation
11. Science
At level 7, minds construct models to test and select alternatives -
and at this level memes are now active and give rise to trial and error
without the selective costs to genetically- and biologically based interaction.
Memes must result in some outcome that can achieve success or not, on the
basis of which they persist over other memes, to be selected at all. The
details of Campbell's schema are not important here, but memes must
arise at some level of information processing complexity, and must
affect outcomes at levels higher than that. And the converse is also true.
Of any cultural process that involves transmission and selection, there
must be memes that "constitute" phemotypes at that level.
Take two examples - a personal attempt to try out a hitherto unlearnt linguistic
form and a scientific theory. When one is learning a first language, syntax
and semantics are underdetermined. A child cannot know, except through
trying out combinations and seeing the results, whether a certain "hypothesis"
about the use, structure and referents of a word, phrase or syntactical
structure is correct. For example, consider how children first learn regular
declensions of noun plurals and regular conjugations of verb tenses, and
are then corrected by more competent speakers when they use a regular form
of an irregular noun or verb - "I swimmed", "he runned",
and so forth. Not only is there an internal selection process going
on through increasing experience of success, but in a formal sense, these
irregular abstract constructs of language are defined through the
role that they play in the trial and error. Of course, language in itself
is not a solitary pursuit, but in individual learning, the trial
and error selection process is (figure 4, cf. Cziko
1995, chapter 11, Ghiselin 1997, chapter 9).
|
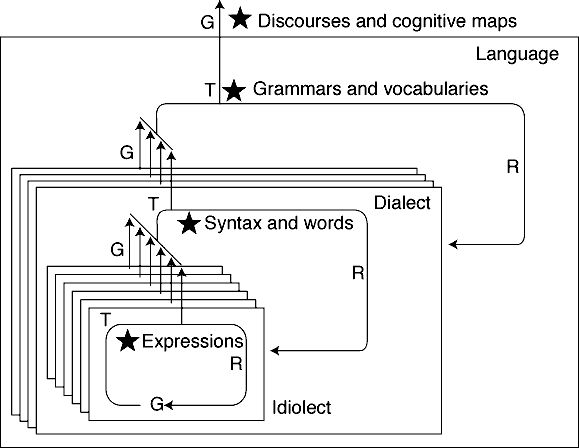 Figure 4. A selective hierarchy
of language, following Ghiselin 1997
Figure 4. A selective hierarchy
of language, following Ghiselin 1997
|
Back to Dawkins' scientific notions: whatever Mendel, or Darwin, or
Newton, internally intended their theories to mean or cover, until the
elements of their theories - supporting data, formalizations, explanatory
rationales and so forth - were published (literally, made public),
discussed, tested, and above all in further scientific work, they weren't
scientific theories as such. What's more, their theories contained memes
of which they were not aware, because they only developed when selection
pressures were applied to them. "Action at a distance", or "survival
of the fittest", became memes so far as they needed to be defended
and supported in the scientific debate when challenged. It is the hallmark
and defining characteristic of science that it is presented and tested
in the public domain of fellow professionals; and this process modifies
theories. Elements of them are quietly or noisily rejected, abandoned or
simply never followed up (Laudan 1977). What
are the memes of a theory, then? They are those elements of it that are
subjected to selection through testing and co-adaptation with the rest
of the theory and of the wider field of the scientific culture (Wilkins
forthcoming). Theories, like Whitman, contain multitudes (and can therefore
contradict themselves), but scientific work consists in large part in exploring
the ramifications of a model, in order to eliminate contradictions, either
in terms of self consistency and coherence, or in terms of agreement with
method and observation.
In the special case of memes in scientific theories, whether or not
Dawkins intended it, it becomes clear that in a general evolutionary model
a replicator is defined by (1) being transmitted intact, and (2) being
subjected to selection. Many things are transmitted in culture that are
not selected, and many things are selected that do not get transmitted.
I conclude that a meme is something (the "smallest" something
you can identify) that gets replicated and selected in culture as a unit
and therefore offer the following short definition of "meme".
A meme is the least unit of sociocultural information relative to
a selection process that has favourable or unfavourable selection bias
that exceeds its endogenous tendency to change.
Exceeding endogenous change simply means that it gets transmitted intact
more than it gets transmitted mutated (Dawkins' condition of fidelity);
in other words, it is more information than noise. Compare this with Williams'
evolutionary gene above.
A single memetic interactive trait, I shall call it the pheme
by analogy with a phene or Mendelian character, is the expression
through some behavioural regularity of a meme at the level of selection.
"Behavioural" here refers to interactive activity at the level
of expression, and so includes mental behaviour, individual behaviour,
group behaviour, and so forth (figure 2). However,
the behaviour must be both causally effective in terms of acquiring
resources and must be in some manner empirically quantifiable. One
question that memeticists should ask of any analysis is "what are
the resources"? A scientific theory may gather resources of researchers'
time, grants and lab space, publication space in the appropriate journals,
and from what I can gather, postgraduate students appear to be a prized
resource. Hull's (1988a, 1988c)
mechanism is that individual scientists seek to acquire credit, either
through innovation, or more probably (since innovation is rare) by inclusion
in a successful research project through citation and extension of results.
An idea in an individual mind will need to gather attention space, memory
resources, and time, in order to out compete other ideas. Without an answer
to the question of what resources?, and they must be measurable,
any memetic account is merely anecdotal.
Sidebar: phenotypes, phemotypes
and classification
A phenotype in biology, and
especially in taxonomy, is the whole array of organismic characters of
an organism, and is distinct from its genotype.
In the taxonomic debate between cladism
and pheneticism described
in Hull's book, the resulting consensus was that there were no privileged
features that are phenetic characters
(Hull 1988c, Ridley 1992)
- anything that can be measured might be used to classify organisms and
reconstruct phylogenetic relationships. This argument applies just as well
to reconstructions and classifications using genetic sequences as it does
to skull shape and the presence or absence of feathers. Some taxonomists
think that the best way to identify species is to make use of non-adaptive
characters on the grounds that these are less likely to be the result of
convergent evolution. The popular belief that it is the entire phenotype
that is the interactor in evolution is, in my view, wrong. Interaction
occurs at the level of the trait, and the "engineering" fitness
of an organism - how well it makes a living overall - is the product of
its various interactive traits. The analogy I wish to make is that memes
result in behavioural regularities that have interactive properties, that
is, which result in social successes and failures. The phemotype
is the array of the phemes that
are the causal outcome of memes in an individual or group. Phemotypes are
the basis for grouping social phenomena, that is, for classifying memetic
ensembles into religions, communities, traditions and programs.
|
Once we have identified a meme in a particular process, though, that
is not the end of the explanatory story. Memetics covers what is common
to the spread of strategies, musical phrases, linguistic practices, ideas
and theories, but it does not exhaust the research. Again the parallel
with genes is instructive. Having identified that there are factors that
are inherited which cause ecologically significant traits, geneticists
moved on to identifying the physical processes and their effects, which
turned out to be extremely complex, with genes that code for traits ranging
from the sorts of proteins that a cell expresses, to genes that regulate
other genes, to genes that contribute to a range of gross morphological
traits at every level of the organismic structure. Identifying memes will
assist in identifying the sorts of cultural phenomena that are functionally
transmitted, but we should expect that these will in turn become causes
of phenomena in another level of analysis.
4 The lineages
and ecologies of culture
Any chain of descent will create a lineage
over time and space if ancestral entities terminate at some stage. Given
that the obverse of a replicator is an interactor, an economic entity that
competes in a formal sense with other interactors for the same resources,
we need to identify the resources, the ecologies, of memes. On the face
of it, memetic resources include neurological and behavioural time and
space, much like a program can tie up the CPU cycles and screen space of
a computer, but they must also extend to the more highly derived necessities
of social life, including such things as credit, currency, and so forth.
Selection processes define not only the replicators (the memes) and the
individuals, but also the active and relevant resources of the ecology
and economy of social life. The interplay between these facets of memetic
evolution leads to successful and persistent lineages, as well as transitory
and unsuccessful ones.
The nature of and reasons for lineages in biological evolution is a
complex matter. Debates about the "Species Problem" and the relationship
between "horizontal", or "non-dimensional" conceptualizations
of species - known generally as biological species concepts - and "vertical"
or "phyletic" conceptualizations of lineages over evolutionary
time - paleontological, evolutionary and phylogenetic species concepts
- have been ongoing since at least the modern synthesis, and indeed is
sometimes held as the defining debate of the modern synthesis of evolution
(Dobzhansky 1937, Mayr
1970, Mayr and Provine 1980). One thing
that is generally agreed upon is that ancestor descendent lineages that
do not regularly recombine ( reticulate
) at a scale on the order of one generation are not species, but are within-species
lineages or something else; for example, what Eigen
(1993) calls quasispecies
in the case of viruses. In other words, species are those largest collections
of biological lineages that do regularly recombine, and which do not themselves
recombine with other lineages.
However, socio-cultural lineages appear to exchange memes on an almost
unconstrained basis. Practices, metaphors, and ideas transfer from one
cultural lineage, culture or tradition to another with consistent ubiquity.
Autochthonous cultures the world over have adopted a range of cultural
artefacts from the dominant western culture, for example, and this pattern
of dominant cultural invasion recurs through recorded and archaeological
history. This reticulate evolutionary pattern is touted as one of the major
disanalogies between cultural and biological evolution. If cultural transmission
is so fluid and free, it is thought, what use is the transfer of biological
to cultural categories and modes of analysis? I shall argue that this objection
rests on an incorrect assumption about biology, founded on our metazoan,
vertebrate, and mammalian prejudices, and that even if it were true, it
would not undermine the memetic enterprise.
Other than accepting the conclusion of this argument from disanalogy
and abandoning memes as useful theoretical entities, there are three alternative
responses to the difficulty it raises for memetics. The first is to shift
ground, and to treat memes epidemiologically; that is, using a metaphor
of disease rather than of evolution. Treating memes as pathogens, and trying
to analyse the vectors and pathogenic dynamics of memes, is a strategy
adopted by Goodenough and Dawkins (1994),
Lynch (1996) and others (cf. Dennett
1995:364-368). "Mind virus" is itself a tolerably good meme.
The second approach is to restrict memetic analysis to domains like the
sciences, where there is a strong selective pressure and a good record
of the spread of scientific ideas, and where lineages tend, within certain
limits and until recently, to be relatively isolated and sui generis.
The third is one that I have not seen explicitly presented but which is
implicit in the work of Hull (1988a, 1988b,
1988c). This is to see that evolution produces
a continuum of results from entirely isolated lineages with the "bridgeless
gaps" so beloved of Mayr resulting from obligate
sexual recombination through to moderately hybridizing lineages like those
of flowering plants and ferns, to regularly recombining lineages - rarely
if ever present in zoological biology but frequent in bacterial phylogeny
and some forms of culture. Each of these approaches has its merits.
The first response, the epidemiological model of memes, takes
into account the rapidity of transmission and mutation (relative to the
observers) of certain varieties of cultural items like vernacular speech
and fashions, and the varieties of strategies they evolve to invade cultural
agents and populations. This view does not, however, obviate the need for
an evolutionary, selectionist, account of the pathogens (cf. Ewald
1994). It's a matter of perspective. Being hosts to so many pathogens,
parasites and symbiots, humans tend to conceptualize disease as sub-generational
and populational events in a relatively short time scale. Nevertheless,
epidemics and pandemics like malaria have evolutionary effects on humans,
resulting in evolutionarily stable genetic equilibria like the maintenance
of sickle cell heterozygosity in regions affected by the transmission of
the malarial parasite Plasmodium falciparum by some species of the
Anopheles maculipennis mosquito group.
But we also often overlook the fact that, from the perspective of the pathogens,
humans and other animal hosts are just another part of their ecosystem,
and our immune systems act as selective environments affecting their
evolution. It's also a matter of degree: viruses, for example, mutate much
more rapidly than the gametes of a metazoan, but this is a difference in
rate rather than kind. There is a bias imposed on us by virtue of being
multicellular, animals, and living at the time scale we do. Evolution has
no such bias. The equations governing (or describing) epidemiology are
re-castings of the Fisherian and Wrightean equations covering (or describing)
natural selection (Cavalli-Sforza
and Feldman 1981), which is what we should expect if we see epidemiological
phenomena from the pathogenic perspective and not from the host's.
What this signifies for memetics is that meme epidemiology needs to clearly
distinguish between human minds and memes as entities that generate their
own separate lineages. More on this shortly. For now, it is worth observing
that meme lineages may not be so obvious as we think, and that cross-lineage
borrowing, which occurs within biology as well as culture, may be less
frequent than seems the case when we view memetic evolution from the perspective
of the host minds.
The second response, the narrow conception, is to restrict the
application of memetic analysis to those cultural phenomena which are phenomenologically
isolated, well recorded, or in which there is a clear selective pressure
such as in technological, scientific or economic change. There is some
merit to this: but it is a heuristic merit rather than a theoretical limitation.
It is a bit like the situation facing early genetics. Genes were, in the
heyday of the Mendelian revolution, theoretical entities that some of a
positivistic bent considered mere instrumental constructs. Positivists
held (and hold) that theoretical constructs have no ontological import.
"Genes" were useful in calculation and modelling, but it was
illegitimate to infer that any such biological entity existed. In retrospect,
this made a metaphysical virtue of a methodological necessity. Once the
science, and ancillary sciences and techniques such as x-ray crystallography,
had developed further, physical genetic entities were discovered and described,
but this was not open to the early Mendelian researchers, and in fact attempts
to describe the physical properties of genes by Weismann and others resulted
in failure and open speculation. Had this positivistic operationalist approach
been taken seriously, the molecular structure of genes might never have
been investigated and discovered.
It might be a good strategy to begin with the more accessible areas
of research like the history of a science, and to develop methods and models
that can be extended later as more problematic cases come within theoretical
reach. In the same manner, biological concepts like "species"
are often developed in relatively clear-cut cases like sexually recombining
animals and plants, and then extended into cases of parthenogenic,
polyploidal, clonal, viral
and other "eccentric" lineages. We must, of course, beware of
biting off more than we can chew, but also of not closing off any further
avenues of research. Memetics, like any science, must evolve gradually.
The third response I'll call the generalist conception.
This is to more broadly characterize biological evolution. As Hull has
said, there is nothing so outlandish that an example of it cannot be found
in biology somewhere. For example, take the cases of reproduction modes
and speciation: memetic evolution appears to be more like the evolution
of plants, which have an estimated 30-40% rate of reticulation, and fungi
and single celled organisms, with monoparental or clonal reproduction,
than it is like zoological evolution with which most non biologists are
familiar. Nearly half of all flowering plants are the result of joined
lineages rather than split lineages, and nearly all fern species. Amoeba
undergo a process called conjugation
where genes are exchanged every few generations, although the rest of the
time they split aparentally. Amazingly, even viral RNA can be exchanged
through crossover of transcriptase in superinfected cells.
Evolution is still cleverer than we are. These idiosyncratic forms of individual,
phenotype, lineage and evolutionary behaviour are nevertheless covered
by the various theories of evolution.
The generalization of evolutionary theory accommodates these "eccentric"
cases just as well as it does the paradigmatic case of animal evolution
on which it was first founded. Natural selection is an emergent process
acting on ecologically interacting replicators, and obligately sexually
reproducing individual organisms are just a special case, even though they
are the most obvious and easily investigated form of evolution for us.
Therefore, it would seem that natural selection can model cultural lineages
even if they do not happen to form tightly integrated populations of recombining
entities. But we need to investigate a posteriori whether any particular
kind of cultural lineage is tightly integrated like an animal species or
not, and not to assume that there is, in fact, such a disanalogy. Some
are, some are not. Of those that are not, selection may be a major cause
of change or it may not. This is an empirical argument, not an axiomatic
derivation. In other words, we should challenge the major premise of the
disanalogy argument in each case. The issue of "quasi-species"
(Eigen 1993) in obligately clonal bacteria and
viruses illustrates this. In principle, and intuitively, every time a clone
or virus generates a mutant, one gets an entirely new strain. We might
expect that strains would indefinitely diverge to fill the entire space
of ecological niches available. In fact what happens is that viruses tend
to cluster about the "wild-type" phenotypic mean (which may not
even exist in a single geneome), forming analogues to recombinant species.
Natural selection accounts for this as well as it does for sexual, biparental
species.
While on the topic of cultural lineages and species, let us consider
the matter of speciation mode.
There is a raging debate about whether sexual species speciate through
allopatry (geographical isolation)
or sympatry (geographical and
ecological coexistence) or some intermediate state ( parapatry
) (Mayr 1970, Gibbons
and Morell 1996).
This resolves to the question whether natural selection, as well as being
responsible for allele frequencies within a breeding population
and for adaptation overall, is also responsible for creating lineages.
Those who, like Mayr, opt for the allopatric and parapatric modes alone,
consider that isolation followed by selectively neutral or decoupled drift
is responsible for speciation, and they tend to see selection as an ancillary
process to speciation rather than a reason for it. Sympatric speciation
is a litmus test of whether one thinks selection is the main or even the
only mechanism of evolution. If species can diverge in the same ecological
territory, it must be because selection causes multiple modes in the one
population, leading eventually to more than one species as the modes chase
different adaptive peaks. If correct, this means that selection can cause
speciation, and the traits of species that differentiate them are adaptive.
If one denies this happens, or that it happens very much, then what differentiates
species can be adaptively neutral or even maladaptive. Nobody presently
thinks that selection can be dispensed with even in cases of speciation
by hybridization of plants, which occurs in a single generation or even
repeatedly ( polyploid speciation),
but the differences in the degree of the attributed importance of selection
points to a deep division in modern evolutionary thought, and this is very
important for memetics if memetics is based on selectively significant
cultural transmits.
Hull (1988c) has addressed similar issues in
scientific evolution. He uses Wright's notion of structured and relatively
insulated breeding populations - called demes
- to characterize research groups. According to him, scientific research
arises first within relatively co-operative demes of scientists - groups
within which more variation is tolerated and selection pressures are less
severe than in the wider discipline. This permits new theories to be developed
and articulated to the stage where they may be able to survive rigorous
criticism by rival groups when published. Intriguingly, Hull does not require
unanimity of opinion within the deme, but merely a desire to increase one's
credit by sharing in the conceptual inclusive fitness of the deme: reputation
is everything in science. Hull's view of the social and conceptual development
of science is a well developed model of conceptual change, one of the most
articulated memetic models yet presented, and it can be extended into other
domains. For example, although Hull does not characterize demonic structure
itself in this way, it can be thought of as an evolutionarily
stable strategy for science to adopt - with a balance between public
debate and private support, conceptual transmission in science is able
to develop a great deal more novelty and yet retain more strongly tested
("more fit") ideas than was possible in the late medieval guild
tradition from which it sprang. Neither a selectively moderate social ecology
nor a naked selective environment where new ideas are never given a chance
could provide the sort of adaptive growth science has achieved. Obviously,
periods of stagnation in a discipline may be due to changes in these parameters
(for example, transmission rates) and connecting them in a model based
on a generalized evolutionary and ecological process is an important first
step to a full memetic analysis of science. Mutatis mutandis, the
same is true of memetic theory in general. If we are to understand how
memes diversify into relatively stable lineages like religious traditions,
the structures within which novelties arise and the selective pressures
to which they are exposed are crucial.
Ecological succession is another analogue of value. It is well
recognised in biogeography that niche occupiers in an ecosystem can often
fend off adaptively superior invaders; the dominance of a given (say) grazer
species in a locale may be due to it being there first rather than its
overall adaptive excellence. We should expect that ecological succession,
which is complex in biology, will be equally or more complex in culture,
but there may be fruitful avenues to explore here. To understand and employ
these analogues, we also need an analogue for "ecology" and "ecosystem"
for a given socio-cultural domain. The ecology of science, for example,
is not nearly as simple as the falsifying crucial experiments that Popperian
cycles propose. The resources for which a social or conceptual structure
competes will usually be fairly broad even if, as in the case of a scientific
idea, storage and processing space and time in an active central nervous
system is the basic resource.
5 Memetic individuals
With the clarifications and mental tools introduced above, we are now
able to ask Hull's question from biology - what is an individual? - in
the memetic context. What is a memetic individual? What is subjected to
selection in culture? What gets "coded for"?
When Juliet bitterly but eloquently complained how Romeo's social relationships
were messing up their love life, she made the interesting observation that
being a Montague, and being the person she loved, were two distinct states
(apparently she had more of an interest in some of Romeo's biological aspects).
The instantiation of the cultural relationship "is a Montague"
in a particular biological organism, denoted by the name Romeo, is a case
where an individual is something other than the sum of his own memes. The
converse argument was given by the idealist philosopher FH Bradley in 1876,
in a landmark essay in ethics, "My Station and its Duties". Here
Bradley wishes to establish that one's social location and relations determine
one's moral responsibilities. To do this, Bradley argues that what we are
as social beings, as moral agents, is determined by the community of which
we are part. An Englishman is not his biology.
In each case, Juliet and Bradley recognize the distinction, often overlooked
by meme enthusiasts, between the biological and the memetic. Memes don't
necessarily make you more biologically fit, nor are they necessarily going
to make you less fit. Memes aren't fit themselves simply because they make
you live healthier lives. Memes are fit only insofar as they are propagated
successfully; forget the effects they have on biology. In Toulmin's
(1972) under appreciated book, he says of biological and cultural evolutionary
processes that they are decoupled. This is not to say that the two
realms do not meet and affect each other, for clearly they do; it is to
say that no matter how you might be able to conceptualize cultural phenomena
in biological terms (socio-biology), you can independently conceptualize
them in social terms (memetics). To lift a phrase of Williams'
(1992) there is a dearth of shared descriptors between the entities
of biology and the entities of culture. Sometimes they may, indeed, be
the same objects or processes, but you have to describe them differently
in each analytic realm.
No end of confusion has been caused by ambiguous specification of the
socio-cultural analogues of phenotype and genotype. An early criticism
of Hull's model of models (Heyes 1988, Tennant
1988) was that scientific memes failed to constitute scientists
in the same way that genes constituted organic individuals. By the same
token, critics argue that selection against memes fails to result in the
death or restricted fertility of scientists, and so natural selection is
an inappropriate way to model scientific theory change. Another recurring
criticism of evolutionary pictures of culture is that unlike organisms
and their genes, cultural agents choose which memes they adopt, and that
they modify their memes in the light of experience. Cultural change would
be Lamarckian, say critics, if it evolved at all. These two attacks on
memetics - the aphenotypic disanalogy and the cultural lamarckism
criticism -- are closely intertwined, and have to be discussed together.
Using the relatively clear case of science as an example, we can see
that any one scientist can adopt differing memetic stances over time. When
Einstein was learning physics, he was probably taught the axiomatic observer
independent Newtonian theories of the day. He later changed his mind about
that. When the physicist Schrödinger got interested in biology, he
entered into the canons and data of a new discipline (for him). Here are
two exemplary cases of memetic acquisition - the replacement or supplementation
of one set of memes with a whole new set. Yet, neither Einstein nor Schrödinger
became different organisms with these memes. Moreover, had either of these
scientists undertaken a non-scientific hobby requiring much technical knowledge
and experience, such as angling, oil painting or cabinet making, the likelihood
is that they would be able to keep them relatively distinct from the professional
standards and methods of physics. So, how can a "physicist" be
caused to come into being by memes, in the same way that a blackbird
is caused by a blackbird genome?
I tacitly answered this question by inserting the word "professional"
above (cf. Wilkins forthcoming). A human
organism who is a member of a culture, language group, class and so forth,
only achieves professional status or competency upon completing some portion
of a "developmental" process of acquiring and exercising the
relevant memes. A scientist, to return to our example, must acquire the
standards, knowledge base and experience of the particular discipline (physics)
and the sub disciplines (for example, high energy physics). There is an
aspect or profile to every scientist that is (a) constituted
by memes, and (b) within the distribution curve of traits exhibited by
the lineage (the phemorph). As a scientist, an organism is distinct
from other cultural profiles he or she instantiates, and the profile is
a separate class of entity from any biological organism of species H.
sapiens or any set of traits that the organism exhibits by virtue of
being a member of the species. Bradley considers the thought experiment
of raising the same biological organism now an Englishman in another society,
or on a desert island. Clearly, says Bradley, not in these words, the properties
of Englishness are derived from the relational and (non-biological) developmental
processes of living in English society and having a locus there. In our
terms, "being English" is a memetic
profile derived from the acquisition of Englishness memes. We therefore
have to be careful of the denotation of the terms "scientist",
"Englishman", "capitalist" and so on. The scientist-as-organism
is a distinctly biological entity, distinct from the cultural
entity scientist-as-Englishman, or scientist-as-capitalist, which depend
for their specification on the memetic profile of the scientist proper,
the scientist-as-professional. In the context of a professional lineage
such as a science, or other intellectual pursuits or social practices such
as accounting or rap dancing, the memetic
individual is the competent member of the lineage, which is developed
by the lineages' professional or cultural properties to create a profile
within the human (biological/neurological) individual organism. With appropriate
adjustments, the notion of a memetic individual can be generalized from
the case study of science into other discernible cultural institutions
and traditions. In the limiting case, where the cohesion of a lineage is
very loose, memetic individuality may be partial or fragmentary (like being
good at using a yoyo), but if memes constitute anything at all,
they constitute the profile of a memetic individual.
Memes, like genes, can only "code for" a norm
of reaction. All cultural as well as biological traits are distributed
over a population curve, with the mean and the mode correlating to the
memetic selection bias. As Hull observes, no two scientists even with identical
theoretical commitments interpret their views exactly the same way, and
it is an oft repeated half joke, half complaint, that there are as many
views in a research program as there are practitioners, sometimes even
more. Neither memes nor genes determine all aspects of the properties of
the entities they constitute. What they do determine are the degrees of
freedom and they bias and constrain the outcomes of the so-constituted
system. In thermodynamic phrasing, they specify the field of states such
memetic systems have a propensity to attain. Do they do this through a
process analogous to the Weismannian Central Dogma, with a one way germ
lineage, or through something analogous to a Lamarckian pangenesis
(Hull 1988c, chapter 12) with the environment
instructing the memes? Is the question even relevant? Cultural and biological
evolution are going to be different in the frequencies of the kinds
of processes they undergo. Perhaps culture does exhibit Lamarckian-style
inheritance through the sort of environmental instruction that has a never,
or rarely, occurs in biological evolution. One outcome of this would be
that variation would come more frequently and more intensively than in
a purely Weismannian process, which has to await random mutation or use
stored variation from earlier mutation in order for selection to have something
on which to operate (Fisher 1930). It would,
however, still be Darwinian evolution even if inheritance
were Lamarckian. Lamarckian evolution, however, is a different sort
of process altogether, driven by perceived need to achieve foreseen outcomes;
(see7). Lamarckian inheritance
is not inconsistent with a Darwinian model of memetic evolution. Lamarckian
evolution would totally demolish the foundations of memetic theory,
and leave us with more traditional forms of cultural analysis. However,
I do not think that the case has been made that memes are even acquired
through instruction in the Lamarckian sense, and refer readers to Cziko's
(1995) discussion for further consideration.
It is my opinion that cultural inheritance is not particularly creative,
and that most "novelty" is in fact the recombination of pre-existing
memes in novel ways: there is little that is new under the sun. Neither
do I think that the rate of creativity rises in times of stress or great
change. I speculate that what changes, and gives the appearance of memetic
novelty, are such factors as selective pressure coefficients, "migration"
rates, and density dependent recombination rates that vary from the "usual"
background rates. Problems can persist for long periods even in the face
of urgent need, and unless the appropriate combination of memes occurs,
more or less at random,
they do not get solved. This takes us back to the question of the demonic
structure of culture - too great an isolation and a more optimal memetic
combination is unlikely; too little isolation, and combinations are unlikely
to be stable enough to undergo selection, and will be swamped. The one
major disanalogy Hull accepts between biological and cultural evolution,
and with which I agree, is in the rate, but not the kind, of reticulate
phylogeny; that is, in the merging and crossover of memes between lineages.
Again this is not so much a difference of kind as of degree. As noted above,
cultural evolution resembles that of bacteria, plants and fungi more than
that of animals - Marvell's "vegetable love" that grows "vaster
than empires" is perhaps the better analogy, although on an evolutionary
time scale, culture is anything but "more slow".
It might help to visualize the analogy I am proposing by giving examples
in a table of the analogues between biological and cultural entities and
processes, mapped to the Hull-Dawkins and Ghiselin-Hull ontologies (Table
1, cf. Ghiselin 1997 for a review).
| Entity (e) or process (p) |
Biological |
Memetic |
Science |
Language |
Economics |
| Interactor (e) |
Phenotype or trait |
Phemotype or Pheme |
Experiment or Observation |
Linguistic behaviour |
Enterprise or transaction |
| Replicator (e) |
Gene |
Meme |
Theory or Hypothesis |
Language element |
Account, resource |
| Character (e) |
Mendelian Trait (phene) |
Pheme |
Method, Result |
Linguistic practice |
Business practice |
| Lineage (e) |
Species, phylum |
Tradition, institution |
Research program |
Regional dialect, language group |
Business, industry |
| Economy (e) |
Ecological system |
Culture |
Disciplinary community |
Language community |
Market sector, fiscal system |
| Reproduction (p) |
Organismic reproduction |
Constitution of new profile |
Degree, training |
Language acquisition |
Establishment of new enterprise |
| Individual (e) |
Organism, kin group, colony |
Memetic individual |
Scientist |
Language speaker |
Economic agent |
| Substrate
(e) |
Organic molecules |
Neural networks
|
| Code (p) |
DNA alphabet |
Semantic |
Semantic and mathematical |
Grammar and vocabulary |
Currency |
| Encoding medium (e) |
Amino acids, proteins |
Neural structures, practices |
Neural structures, journals, books, institutions, disciplinary
associations |
Neural structures, written material, recordings, etc. |
Neural structures, computers, books, ledgers, receipts,
bank files, etc. |
Table 1 Examples of general evolutionary analogues (e=entity,
p=process)
It must again be stressed that these are only
some of a number of possible sets of analogues, and what functions as an
interactor at one level may function as a replicator at another, so that
a pheme for one level may be a meme at another, and vice versa.
6
Conclusion and prospectus
6.1 Conclusions: My conclusions are
as follows. Memes are those units of transmitted information that are subject
to selection biases at a given level of hierarchical organization of culture.
Unlike genes, they
are not instantiated in any exclusive kind of physical array or system,
although at base they happen to be stored in and expressed from neurological
structures. Many memes
reside as neural net structures in the central nervous systems of humans,
but many also emerge at a higher cultural level. All memes have neural
substrates, but not all are encoded in those substrates. Memes are also
situated in a variety of larger semantic structures, behavioural regularities,
and cultural substrates. They are identified in virtue of their selective
roles. Memes must be expressed in a cultural ecology in order to be selected,
but it is the class of behaviours rather than the behaviours themselves
that are memes. Memes do not control behaviour (including mental behaviour)
rigidly, but bias and constrain it to a norm of reaction. Memes are the
replicators of cultural evolution and the structures that bear the cultural
properties they express as are the interactors, in the language of the
Hull-Dawkins Distinction. They are, as Hull once entitled a paper (1987),
genealogical actors in ecological roles. Packages of memetic interactive
properties - phemes - constitute the phemotype of memetic individuals,
or memetic profiles, that are not coextensive with the descriptors of the
biological individuals in which they are instantiated, and cultural evolution
is neither identical to nor derived from biological evolution. Memetic
inheritance may be, but probably isn't, analogous to Lamarckian inheritance,
but in any event, memetic evolution is Darwinian. Memes form ancestor descendent
chains of populations that ramify and reticulate with frequencies differing
from biological phylogeny, but the differences appear to be within the
extremes of the parameters of biology. The models developed for biological
evolution and ecology need to be understood more broadly than just vertebrate
animal evolution and applied as they are suited to culture, in order to
determine the general evolutionary properties of both domains.
6.2 Methods: The methodological future
of memetics lies in the use of techniques drawn from information theory,
modern taxonomy and computer science. By interpreting memes as messages,
we can make use of Shannon-Weaver entropy (cf. Brooks
and Wiley 1988), cladistics (Wiley, et al
1991), and connectionist mathematics as implemented in the Artificial
Life program (Holland 1995), but as each of these requires quantification
of the data or input, it is important to be able to commensurably map the
memetic elements and to measure the selection biases they undergo. If,
as I have argued, a meme exists in virtue of biased transmission rates,
then there is no smooth reduction of memetic structures from cultural behaviour
to atomic memes, just as there is no smooth reduction from phenotypic traits
to single genes. The researcher seeking to explain a singular historical
shift of memetic frequencies must iteratively refine the data until it
becomes clear what is being transmitted at a level and how it is being
expressed. The problem of classification lies at two ends of the scale
- identifying cultural traditions as they exist now and over time, and
identifying elements of those traditions as they persist and recombine.
Although this sounds subjective, it need not be. Behavioural regularities
indicate that something objective has been spread, and even if the underlying
memes cannot be formulated in some universal logical language, the structure
of memes can still be identified in the same way as Mendelian genes and
molecular sequences, through the use of consensus maps and by noting when
their absence or presence makes a difference.
The analytic tools available to us are legion,
having been developed in biology over a century or more. They include Wagner
groundplan methods (unrooted character similarity trees based on presence-absence
matrices), Hamming Distance measures (the sum of the number of simple differences
between two memes),
cladistic reconstruction using parsimony methods, and pattern recognition methods that use neural networks and other
connectionist models. Eventually, these and other methods will no doubt
be incorporated into a body of canonical techniques within the memetic
enterprise, and I expect they will be generalized as methods applicable
in a range of social disciplines. The methods being developed for complex
adaptive systems theory at the Santa Fe Institute are also likely to become
important for memetics as well as other research dealing with complex adaptive
phenomena (Casti 1994). However, if we lack clarity
on the core ontology of a selection process in culture, the data to which
we could apply these methods will be subjective and the explanations that
we derive from them are in danger of being entirely vacuous.
Acknowledgements
I am indebted to Mario Vaneechoutte and Mark
Mills for critical comments on an earlier draft of this paper, and the
referees for subjecting the submission to strong selective pressures on
style and content. I must also thank the late David Rindos for encouragement.
Major stylistic improvements are due to the attention of Vladimir Brusic,
though he is not to blame for any remaining infelicities.
Glossary
of technical terms used
- allele
- An alternative gene at a particular locus.
- allopatry
- The state of populations living apart from each other (literally "other
homeland"). Hence, allopatric speciation.
- Central Dogma
- Weismann's hypothesis that gametes ("germ
cells") are passed on independently to what happens to the organism's
body.
- cladistics
- Taxonomic classification that reconstructs the order of the appearance
of evolutionary novelties from their present distribution. Also known as
cladism.
- cladogenesis
- The origination of a new species by the splitting
of a single lineage into two. See speciation.
- co-adaptation
- The process of local adaptation of genes and their
effects to each other, so that they function well as a unit.
- conjugation
- In some single celled organisms, the occasional exchange of genes
in otherwise aparentally reproducing organisms.
- deme
- A small population that is relatively isolated from the larger species
or tradition and has its own distinct genetic or memetic characteristics.
- emergentism
- The metaphysical position that properties arise from the relations
of objects that are not properties of the objects themselves.
- epistasis
- Linkages in the effects of genes, such that a single
gene may affect many traits or a single trait may be affected by many genes.
- evolutionary
gene
- "[A]ny inherited information for which there is a favorable
or unfavorable selection bias equal to several
or many times its rate of endogenous change" (Williams
1966: 25)
- evolutionarily
stable strategy
- A strategy coded for by genes or memes
that does better when interacting with copies of itself than alternative
strategies do, and which will tend to become the sole or dominant strategy
of a population that will not then be susceptible to invasion by other
strategies.
- facultative
- Of an organism, the possibility to adopt variant lifestyles, one of
which is the norm.
- founder effect
- The evolution of a new lineage based on the
sampling errors of the small starting population, which may be of different
proportions to the original populations.
- gamete
- The sex cell of each parent, which recombines to produce the zygote,
such as sperm and egg, or spore.
- gene
- The fundamental physical unit of heredity that transmits information
from one cell to another and thus to successive generations.
- Mendelian gene: a unit of heredity
that causes a single phenotypic
character or trait.
- Molecular gene: a sequence of nucleotides
(DNA, tRNA or rRNA) that functions as a unit during transcription and which
is transmitted whole.
- Evolutionary gene: see evolutionary
gene.
- genet
- All the clonal entities that share a genotype.
- genome
- The complete complement of the genetic material in a cell or carried
by an individual.
- genotype
- The genetic constitution of an individual,
often referring to genetic basis of some particular characters.
- heterozygosity
- Carrying two different alleles from each parent
at a locus.
- individual
- A relatively well bounded and functionally coherent system comprised
of components and their relationships. The formal opposite of a class or
universal type.
- interactor
- "[A]n entity that interacts as a cohesive whole with its environment
in such a way that this interaction causes replication to be differential"
(Hull 1988c: 408).
- lineage
- "[A]n entity that persists indefinitely through time either
in the same or an altered state as a result of replication" (Hull
1988c: 409).
- meme
- The least unit of socio-cultural information relative to a selection
process that has favourable or unfavourable selection
bias that exceeds its endogenous tendency to change.
- memetic profile
- The array of phemes that constitutes
a memetic individual.
- memetic individual
- A competent member of a memetic cultural lineage,
which is developed by the lineages' professional or cultural properties
to create a memetic profile
within the human (biological/neurological) individual organism.
- methodological
individualism
- The philosophical belief that collectives and their properties are
just the sum of the individuals and their properties
that comprise them, especially in social and historical explanation.
- mnemone
- Donald Campbell's term for a conceptual replicator,
roughly equivalent to meme.
- norm of reaction
- The distribution curve of the phenotypic effects
of a gene in a population.
- obligate
- Of a parasite, the forced mode of living or the necessary host. In
general, the lifestyle that an organism is forced to adopt.
- pangenesis
- Darwin's theory of heredity, a use
inheritance view, in which parts of the body were supposed to throw
off "gemmules" that were carried to the reproductive organs,
and which carried information about the body's experience to the next generation.
This view was discredited by August Weismann in the 1880s.
- panselectionism
- The view that all characters of an organism have an adaptive reason
for evolving.
- parapatry
- The state of living in adjacent regions with or without some overlap
("bordering homeland"). Hence, parapatric speciation.
- parthenogeny
- Asexual reproduction through unfertilized eggs of a lineage
that evolved from a sexually reproducing ancestral state. Adj. parthenogenic.
- pheme
- A single memetic interactive trait which is the expression through
some behavioural regularity of a meme at the level
of selection. It is the least type of selectively biased behaviour relative
to a culture.
- phemorph
- The normal distribution curve of traits exhibited by a cultural lineage.
- phemotype
- The array of the phemes that are the causal outcome
of memes in an individual
or group.
- phene
- A Mendelian character or trait. Adj. phenetic.
- pheneticism
- Taxonomy based on the groupings of phenes, without
respect to evolutionary lineages.
- phenotype
- The observable features of an organism, which develop according to
its genetic code (genotype). Adj. phenotypic.
- pleiotropy
- The state in which one gene affects two or more
phenotypic traits not otherwise
directly related.
- polyploidy
- The fusion of three or more complete sets of chromosomes, sometimes
from distinct species, usually in plants.
- quasi-species
- Manfred Eigen's term for clusters of related clonal organisms that
mimic species in the way they remain similar.
- ramet
- An entity that is one of a number of genetically identical organisms.
- random drift
- Sewall Wright's model of random allele frequency changes in small populations
without the operation of natural selection.
- reductionism
- Explanatory reductionism: the philosophical doctrine that a
complete explanation of a complex whole is given by enumeration of the
components of that whole and their properties, especially in scientific
explanation.
- Genetic reductionism: a form of explanatory
reductionism that has developed into a research program, which holds that
all phenotypic properties of
organisms and their evolution can be understood in terms of genes
and their fitness levels.
- replicator
- "[A]n entity that passes on its structure largely intact through
successive replications" (Hull 1988c:
408).
- reticulation
- The recombination of distinct phylogenetic lineages.
See speciation.
- selection
- "[A] process in which the differential extinction and proliferation
of interactors cause the differential perpetuation
of the relevant replicators" (Hull
1988c: 409).
- spandrel
- Gould's and Lewontin's (1979) term
for a trait or structure that is a necessary by-product of some other adaptive
feature, and which is not therefore explained in terms of selection
in favour of it.
- speciation
- The process of the evolution of a new species, through splitting from
an existing species ( cladogenesis
). Also used of hybridization of existing species to form a third ( reticulation
).
- species
- The largest collections of (sexually reproducing) biological lineages
that do regularly recombine, and which do not themselves recombine with
other lineages.
- sympatry
- The state of coexisting in the same region ("same homeland").
Hence, sympatric speciation.
- use-inheritance
- Also called soft inheritance (Mayr 1982:
691). A view held by Darwin (Origin chapter V) that "use ...
strengthens and enlarges certain parts and disuse diminishes them, and
that such modifications are inherited". Sometimes called "Lamarckian
inheritance", although Lamarck was not the first to propose this view,
which is a folk belief of long standing. In Darwin's theory of pangenesis,
use-inheritance is responsible for modifying the frequency and novelty
of variation, and therefore evolution. Fisher (1930)
discusses the problems of use-inheritance for Darwinian evolution, and
the arguments against it.
Notes
Technical
terms are linked to definitions in the glossary
and discussed, where relevant, in the body of the paper.
That
is, it represents something causally significant in the theory, like an
electron.
An
analogy between two domains depends on a common etiology. A metaphor can
only suggest limited and vague similarities. For instance, the term "analogy"
in phylogeny refers to convergently evolved traits through adaptation to
similar conditions of life. If selection of culture is an analogy, it has
theoretical weight; if a metaphor, we can abandon or modify it when we
find it difficult to apply.
And
Dawkins is a lot of a panselectionist. Panselectionists tend to
find selective explanations for every feature of organisms, for instance
asking what the adaptive significance of the human chin is, when chins
are just the result of two growth fields interacting.
By
"evolution" I mean the generation of variations and adaptation.
I am not committing myself to the claim that natural selection is, or is
not, responsible for new species through cladogenesis.
Weismann
was the first researcher to show that gametes
(he called them "germ cells") are passed on independently of
what happens to the organism's body. His Central Dogma, as it became known,
was an attack on use-inheritance,
the time honoured belief (shared by Darwin among others, cf. Mayr
1982: 687-694, Richards, 1992: 172) that
those features of an organism that are most needed and most used will be
more frequently inherited. Even after the period of the development of
Mendelian genetics, the Central Dogma was not universally accepted by biologists
until the middle of this century and the molecular revolution in genetics.
The
term "Lamarckism" is very ambiguous in the way it is used in
biology. Much historical injustice to Lamarck was done by those who called
themselves "neo-Lamarckians" in the late nineteenth century.
Only two sense of "Lamarckism" are relevant to this paper: use-inheritance
(see previous note), also known as the inheritance of acquired characteristics;
and the belief that novelties evolve to meet the needs (sometimes
interpreted as the desires) of the organisms when their environment changes.
A third form often and more historically accurately referred to is that
evolution is progressing to some form of perfection. The charge that evolution
of culture is Lamarckian (cf. Gould 1993: 216,
1997: 222) generally refers to use-inheritance (for rebuttals, see Hull
1988: 452-457, Cziko 1995).
More
recently, Ghiselin, with whom Hull collaborated on the so-called individuality
thesis, has reviewed the arguments both have made against the "entification"
of replicators, including noting that genes, in the population (or Mendelian)
genetics sense, can include deletions, that is, the loss
of a sequence can have population level effects. In his view, this is a
reductio ad absurdum of Dawkins' genetic reductionism (Ghiselin
1997: 143-148).
But
it is worth noting that they can be the product of the properties
of genes. In algebra, sums are linear and additive, while products are
non-linear and multiplicative. Non-linear products can be extremely complex,
and are so-called because their functions do not result in a straight line
on log log graphs.
The Kolmogorov theorem shows that between any
two arbitrarily large sets there can be at least one mapping so long as
there is an intermediate set of links, an important result for connectionist
systems research. These Kolmogorov mappings are non-linear products, and
applied to genes and memes the theorem means that while there is no simple
reduction from interactive characters to genes/memes a mapping relation
does exist, even if it is extremely complex.
See
Rosenberg's 1994 discussion and clarification
of the Hullian entities as they apply to biological processes, and also
his discussion of Hull's problem of reductionism. See also Ghiselin
1997.
To
avoid unnecessary confusion, it should be noted that any number of hierarchies
can occur in a single domain like biology or culture. There may be a genetic
hierarchy leading to species, but there may also be a hierarchy leading
to ecological patterns, and these need not be the same hierarchy. (Eldredge
1989) This figure and the following figures are not intended to represent
the hierarchy of that domain, but only one of a possible many.
However,
this does not deny that a selection process based on a mental model of
the states of affairs can occur in an individual mind. In terms of figure
2, the memes here are the entities that are selected in a mental environment,
that is, what makes sense to the thinker, and which when expressed will
become memes in a higher level process [cf. Dennett
1995, chapter 12].
One
possible exception to this bifurcation is when a meme is a spandrel,
that is, a by-product of some other meme that is selectively
favoured, and with which it inevitably must be transmitted (Gould
and Lewontin 1979, Gould 1997).
This
resembles the Marxist-Leninist doctrine of false consciousness in some
ways, but with no presumption that any part of society will instantiate
a "true consciousness" of social reality.
Discussion
in Cziko 1995: 140-149.
For
example, a practice may spread just because it is selectively neutral.
Junk DNA can be transmitted because it does not generate products that
are "visible" to selection. Kimura's theory of neutral DNA shows
that biassed transmission without selective advantage is a biological reality.
And any trait at the interactor level can be selected for or against, in
a sense, even if it is not heritable. A blacksmith may do well biologically
because of his strength, but that will not (necessarily) be bequeathed
to his progeny.
As
we shall see below, this is not a complete picture; but it captures the
initial intuitions of taxonomists.
Interestingly,
identifying the five or six distinct, but hitherto unrecognized (cryptic),
species of A. maculipennis was accomplished through analysis of
their being vectors for P. falciparum or not (Cain
1954).
I
am of the philosophical school of thought that laws and equations merely
describe dynamic patterns rather than govern phenomena (instrumentalism),
but that is not germane to this essay. See Rosenberg
1994 for a discussion of instrumentalism and biology.
Because
the resources required are rather significant, which is an interesting
memetic point.
This
is not a mistake, despite its contradiction to textbook biology. The result
was announced in a recent paper (Boerlijst,
Bonhoeffer and Nowak 1996).
It
is worth remembering that Darwinian, or rather synthetic, evolution, is
modelled by a number of theories (common descent, natural selection, sexual
selection, biogeographic distribution, Mendelian genetics), and should
not be considered a single theory on its own.
Allopatry
is the state of populations living apart from each other (literally "other
homeland"), sympatry that of coexisting in the same region ("same
homeland"), and parapatry of living in adjacent regions with or without
some overlap ("bordering homeland"). When speciation occurs in
these states, then it is called allopatric speciation, and so forth. Many
now think that parapatric speciation occurs, but the frequency is still
at issue.
Dobzhansky's
concept [1937] of "co-adaptation"
(a term initially used by Darwin, I believe) is significant at this point
of the argument. If memes at first arise within restricted domains, all
they need at this early stage to adapt to are the other memes within
the deme, or some part of it. Co-adapted memes may play a role in storing
memetic variation until the fitness of variants rises to significant levels.
Moreover, it would explain why apparently maladaptive memes can persist
in a tradition. In the end, the final selection event is extinction, and
maladaptive memes may persist because they are fitter within a tradition
than on their own, even if they make the lineage phemotype less adaptive
overall.
As an example, consider the meme of common
property in the fideist traditions such as the Mennonites and the Shakers.
This meme was a core belief and tradition in these communities, and was
well adapted to the rest of the memes shared by the communities, but it
was maladaptive during the most aggressive period of the rise of modern
industrial capitalism, and these communities eventually faded away.
An
early shot at proto-sociobiological notions, although I think Bradley underestimates
how much biology affects one's place in a culture. Imagine the role
of a Chinese child growing up in Victorian England.
Random
with respect to the selection process, that is.
Phonemes,
words, syntactical structures, etc.
Funding
for research is peer reviewed, even if it derives from governments, and
so the primary economic resource is standing in that community, cf. Hull
1988c.
Supporting
physical entities of the code.
Genes
also include and require RNA of various kinds, so even this generalization
is not absolute.
At
least, until artificial intelligences become a reality.
Interestingly,
Hamming Distance was first formulated in the context of information theory,
and adapted for use in genetics after the molecular revolution. See Gabora
forthcoming for a treatment of Hamming Distance in memetics, but one
that I think is overly reductionist.
Cladistic
reconstructions rely on an absence of reticulation, and so should be used
with care in the memetic context. Cf. Wiley et
al 1991.
References
- Beatty, J: 1987. Dobzhansky and Drift: Facts,
Values and Chance in Evolutionary Biology. In The Probabilistic Revolution:
Volume 2: Ideas in the Sciences, L Krüger, G Gigerenzer and MS
Morgan, eds., MIT Press, Cambridge MA, 1987
- Boerlijst, MC, S Bonhoeffer, and MA
Nowak: 1996. Viral Quasispecies and Recombination. Proceedings of the
Royal Society of London, Series B, 263: 1577-1584
- Bradley, FH: 1876. Ethical Studies,
Cambridge University Press, Cambridge UK
- Brooks, DR and EO Wiley: 1988. Evolution
as Entropy: Toward a Unified Theory of Biology, University of Chicago
Press, Chicago IL, second edition
- Buss, L: 1987. The Evolution of Individuality,
Princeton University Press, Princeton
- Cain, AJ: 1954. Animal Species and Their
Evolution, Hutchinson's University Library, second edition 1963
- Campbell, DT: 1960. Blind Variation and
Selective Retention in Creative Thought as in Other Knowledge Processes.
In Radnitzky, G and WW Bartley III (eds.), Evolutionary Epistemology,
Rationality, and the Sociology of Knowledge, Open Court, La Salle IL,
1987
- Campbell, DT: 1974. Evolutionary Epistemology.
In Radnitzky, G and WW Bartley III (eds.), Evolutionary Epistemology,
Rationality, and the Sociology of Knowledge, Open Court, La Salle IL,
1987
- Campbell, DT: 1987. Selection Theory and
the Sociology of Scientific Validity. In Callebaut, W and R Pinxten eds,
Evolutionary Epistemology, a Multiparadigm Approach, D Reidel, 1987
- Campbell, DT: 1988. A General `Selection
Theory' as Implemented in Biological Evolution and in Social Belief-transmission-with-modification
in Science. Biology and Philosophy 3: 413-463
- Casti, JL: 1994. Complexification: Explaining
a Paradoxical World Through the Science of Surprise, Abacus/Little,
Brown, London
- Cavalli-Sforza, LL and MW
Feldman: 1981. Cultural Transmission and Evolution: A Quantitative Approach,
Princeton University Press, Princeton NJ
- Cziko, G: 1995. Without Miracles: Universal
Selection Theory and the Second Darwinian Revolution, MIT Press, Cambridge
MA
- Darwin, CR: 1872. The Descent of Man, and
Selection in Relation to Sex, John Murray, London
- Dawkins, R: 1977. The Selfish Gene,
Oxford University Press, Oxford UK (1989 edition)
- Dawkins, R: 1982. The Extended Phenotype:
The Long Reach of the Gene, Oxford University Press, Oxford UK, revised
1989
- Dennett, D: 1995. Darwin's Dangerous Idea:
Evolution and the Meanings of Life, Allen Lane Press, London
- Desmond, A: 1989. The Politics of Evolution,
University of Chicago Press, Chicago IL
- Dewey, J: 1909. The Influence of Darwin on
Philosophy. In Appleton P ed., Darwin: A Norton Critical Edition,
Norton, New York, 1970
- Dobzhansky, T: 1937. Genetics and the
Origin of Species, 3rd edition 1951, Columbia University Press, New
York
- Eigen, M: 1993. Viral Quasispecies. Scientific
American, July 1993: 32-39
- Eldredge, N: 1989. Macroevolutionary
Dynamics: Species, Niches, and Adaptive Peaks, McGraw-Hill, New York
- Ewald, PW: 1994. Evolution of Infectious
Disease, Oxford University Press, Oxford UK
- Fisher, RA: 1930. The Genetical Theory
of Natural Selection, Clarendon Press, Oxford UK (rev. ed. Dover, New
York, 1958)
- Gabora,
L: forthcoming. A Day in the Life of a Meme. Philosophica
- Ghiselin, MT: 1987. Bioeconomics and
the Metaphysics of Selection. Journal of Social and Biological Structures
10: 361-369
- Ghiselin, MT: 1997. Metaphysics and the
Origin of Species, State University of New York Press, Albany NY
- Gibbons, A and V Morell: 1996. On
the Many Origins of Species. Special News Report, Science 273: 1496-1502
- Goodenough, OR and R Dawkins:
1994. The "St Jude" Mind Virus. Nature 371: 24
- Gould, SJ: 1981. The Mismeasure of Man,
Norton, New York
- Gould, SJ: 1993. More Light on Leaves. In Eight
Little Piggies: More Reflections on Natural History, Norton, New York
- Gould, SJ: 1997. The Exaptive Excellence of
Spandrels as a Term and Prototype. Proc Natl Acad Sci, USA 94: 10750-10755
- Gould, SJ and RC Lewontin: 1979. The
Spandrels of San Marco and the Panglossion Paradigm: A Critique of the
Adaptationist Programme, Proc R Soc Lond B 205: 581-598
- Heyes, C: 1988. Are Scientists Agents in Scientific
Change? Biology and Philosophy 3: 194-199
- Holland J: 1995. Hidden Order: How Adaptation
Builds Complexity, Helix Books/Addison-Wesley, Reading MA
- Hull, DL: 1974. The Philosophy of Biological
Science, Prentice-Hall, Reading MA
- Hull, DL: 1987. Genealogical Actors in Ecological
Roles. Biology and Philosophy 2: 168-184, reprinted in The Metaphysics
of Evolution by DL Hull, State University of New York Press, Albany
NY, 1989
- Hull, DL: 1980. Individuality and Selection.
Annual Review of Ecology and Systematics 11: 311-332
- Hull, DL: 1988a. A Mechanism and its Metaphysics:
An Evolutionary Account of the Social and Conceptual Development of Science.
Biology and Philosophy 3:125-155
- Hull, DL: 1988b. A Period of Development: A
Response. Biology and Philosophy 3: 241-263
- Hull, DL: 1988c. Science as a Process: An
Evolutionary Account of the Social and Conceptual Development of Science,
University of Chicago Press, Chicago IL
- Hull, DL: 1992. Individual. In Keywords in
Evolutionary Biology, ed. EF Keller, Harvard University Press, Cambridge
MA
- Kuhn, TS: 1962. The Structure of Scientific
Revolutions, second edition 1970, University of Chicago Press, Chicago
IL
- Laudan, L: 1977. Progress and Its Problems,
University of California Press, Berkeley CA
- Lynch, A: 1996. Thought Contagion: How Belief
Spreads Through Society, Basic Books
- Mayr, E: 1970. Populations, Species and Evolution,
Harvard University Press, Cambridge MA
- Mayr, E and W Provine eds.: 1980. The
Evolutionary Synthesis: Perspectives on the Unification of Biology,
Harvard University Press, Cambridge MA
- Nagel, E: 1961. The Structure of Science:
Problems in the Logic of Scientific Explanation, Routledge and Kegan
Paul, London
- Plotkin, HC: 1994. The Nature of Knowledge:
Concerning Adaptations, Instinct and the Evolution of Intelligence,
Penguin, London
- Ridley, M: 1986. Evolution and Classification:
The Reformation of Cladism, Longman, New York.
- Rosenberg, A: 1994. Instrumental Biology,
or the Disunity of Science, University of Chicago Press, Chicago IL
- Suppe, F: 1989. The Semantic Conception
of Theories and Scientific Realism, University of Illinois Press, Chicago
IL
- Tennant, N: 1988. Theories, Concepts and
Rationality in an Evolutionary Account of Science. Biology and Philosophy,
3: 224-231
- Toulmin, S: 1972. Human Understanding,
Cambridge University Press, Cambridge UK
- van Fraassen, B: 1980. The Scientific
Image, Clarendon, London UK
- Wiley, EO, D Seigel-Causey, DR Brooks,
and VA Funk: 1991. The Compleat Cladist: A Primer of Phylogenetic Procedures,
University of Kansas, Museum of Natural History, Lawrence, KA. Special
Publication Number 19.
- Wilkins,
J: forthcoming. The Evolutionary Structure of Scientific Theories.
Biology and Philosophy
- Williams, GC: 1966. Adaptation and Natural
Selection: A Critique of Some Current Evolutionary Thought, Princeton
University Press, Princeton NJ
- Williams, GC: 1992. Natural Selection:
Domains, Levels, and Challenges, Oxford University Press, Oxford UK
- Wynne-Edwards, VO: 1962. Animal
Dispersion in Relation to Social Behaviour, Oliver and Boyd, London
Back
to Volume 2 - Issue 1